Л кування при остеохондроз
РГ6 од
- 1 AHR 1996
ХАРЮВСЬКИЙ ДЕРЖАбНИЙ МЕДИЧНИЙ УН1ВЕРСИТЕТ
На правах ругапису
КНЯЗеВА Марина Владиславовна
МЕТАБОЛ1ЧН1 АСПЕКТИ 3MIH У ТКАНИНАХ ОРГАН1В:К1СТКОВО-СУГЛОБОВО! I СЕРЦЕВО-СУДИНН01 СИСТЕМ ПРИ CTPECI
03.00.04 - б!ох!м1я з бгаг.опчнпх наук
Автореферат
дисертацД на здобуггя наукового ступеня доктора бгалопчних наук
Харк1в- 1995
Дисертащею е рукопис
Робота виконана в Харшському 1нститут1 удосконалення ж'карш
Науков! консультанти: доктор бюлопчних наук, професор Панченко Нша 1ван1вна доктор б!олОтних наук, п.н.с. Тимошенко Ольга Павл!вна
Оф1ц1йн1 опоненти:
доктор бюлопчних наук, професор Луговий Володимир Йосилович
доктор бюлопчних наук Перський бвген Ефро!мович ,
доктор медичних наук Кульчицький Олег Костянтинович
Пров1дна орган!зац1я - Ки1вський наукопо-досл!дний 1нститут ортопед!! та травматолоп'Т
Захист вщбудеться "11" синя 1996 р. на засщанн! спец!ал!зовано? ради Д 021.36.01 в ¡карювському медичному ун!верситет) в 14-00, 310022, Харк1в, пр.Лен1на,4, ауд. N1
3 дисертац1ею можна ознайомитися в ЦентральнШ науковЮ 61бл1отец! Харкшського медичного ун!верситета
Автореферат роз!сланий "11" грудня 1995 р.
Вчений секрётар спец!ал!зовано! ради кандидат бюлопчних наук
Жубркова Л .О.
Актуальн!сть проблеми
Вивнення метабол1зму i його регуляцИ у тканинах орган!в KicT-ково-суглобовоТ i серцево-судинноТ систем при р!зних BapiaHTax стресу веде до б1льш глибокого розумЫня б1олоМчних механ!зм!в виникнення у них дистроф!чно-деструктивних змш, а також причин сполучення в одному орган1зм1 двох i б!льш захворювань (Горизонтов П.Д., 1976; Парша G.B., Кал!ман П.А., 1978; Тимошенко О.П., 1980; Пан1н Л.6., 1983; Меерсон Ф.З., 1986; Б1ленко М.В., 1989, та ¡н). Сполучення остеохондрозу хребту з !шем!чною хворобою серця (IXC) I гострим .¡нфарктом М10карда (Г1М) с предметом окре-мого вивчення (Froment et Gonin, 1956; Гордон I.E., 1966; Одинцова Т.Н., 1978; Гордон 1.Б., Баранова М.Н., 1984; Алелеков Д.О. з ствавт., 1984; Авак1мян Г.А., 1981; 1ванова Л .А. з сп!вавт., 1984; Поворознюк В.В., 1987 та 1н). Оск!льки у виникненн! кожного з цих захворювань важливе значения мае стрес (Коваленко G.A., Гуро-вський Н.Н., 1980; Чьорнобай Л.В., 1983; Меерсон Ф.З., 1981; 1983; 1966; 1988; Тимошенко О.П., 1980-1990 та 1н.), проблему ix сполучення у одному оргажзм! можна розглядати з позиц/й теорм загального адаптацшного синдрому (Селье Г., 1960-1986). В!домо, що при стрес! резистентн!сть орган1зму е насл!дком 1нтегральноТ роботи велико! к1лькост! функц!ональних систем, що мають caoî особливост1, i д!ючих на р1зних ршнях як едини'й злагоджений механ1зм (Селье Г.,1972;Анох1н П.К., 1975;Судаков К.В., 1984; Розанов О.Я. та ÏH., 1985), у д!яльност! якого м'юце тканин к1стково-суглобово! системи до кЫця не визначено. Анал1з даних л!тератури дае основу для вивчення рол! тканин к!стково-сугло-бово! системи у 1нтегральн!й в1дпов!д| оргажзму на стресов! впливи, poni локальних зм1н у тканинах хребту на метаболизм тканин внутр!шн1х орган1в, а також пошуку загальних метаболпних ланок ¡HTerpauiï м!ж р!зними тканинами. Необх1дшсть комплексного вивчення у тканинах особливостей метабол1зму пов'язана з тим,
1
що у тепер!шн1й час неможливо видтити одну найб!льш 1стотну б!осистему, декомпенсация пко'1 при стресових станах визначае виникнення дистроф!чно-деструктивних змш.
Мета досл!дження - установити особливосп метаболтних реакц1й у тканинах оргажв к!стково-суглобово! I серцево-судинно? систем при р!зних вар&нтах стресу. З'ясувати роль порушень ¡н-теграцн обмЫних процес!в м1ж тканинами як можливу причину виникнення латолог!чних зм!н в к!стково-суглобоый та серцево-судинжй системах при стрес!.
Зпдно з метою досл1дження були поставлен! так! задач!:
1. Установити характер зм!н показнию'в вуглеводного, л!п!д-ного, минерального обм1н1в, вм!сту нукле!нових кислот, а такрж компонента орган!чного матриксу сполучно! тканини у серц1, су-динах, т!лах хребц,1в, м!жхребцевих дисках, компактно'1 частини
■ стегнових к1сток I деяких показникш крое! 3- I 12-м1сячних б!лих щур|а-самц1в при стрес! - 30-добовоТ ппок1нези.
2. Виявити особливост! структурно-метабол!чних зм)н у тканинах орган)в к1.стково-суглобово1, серцево-судинно! систем I у кров! 3- й 12-шсячних щур(в при реадапгацН до нормальних_умов на 1, 3, 5 добу п!сля зняття 30-добового обмеження рух!вно? активност!.
3. Визначити еплив ШдивШуально-типолоПчних особливостей орган1зму б!лих щур!в на характер б1ох!м!чних змж у хребцях, М1жхребцевих дисках, стегнових к!стках, серц!, судинах I кров! при 30-денн!й г1пок1нез11 I реадаптацИ.
4. Установити особливост) змЫ структурно-метабол1чних параметр!в тканин орган!в к!стково-суглобово1 I серцево-судинноТ систем у екслеримент) на модел! дистроф!чного процесу у хребт!, виконаного оперативним шляхом ("експериментальний остеохондроз").
5. Визначити можлив! ланки (нтеграцП б(ох(м(чних процес|'в м1ж тканинами орган1в к1стково-суглобово! I серцево-судинно!' систем.
6. Установити особливост1 метаболЬму у печ!нц1 експериментальних тварин при 30-добово7 г1пок1неэ11, реадаптаи'Т 1 "екс пери ментальному остеохондроз!".
7. Провести пор1вняльний анал1з б1ох1м!чних характеристик тканин оргажв к1стково-суглобовоГ ) серцево-судинноГ систем при 30-добовоГ ппок(нез!Т, реадаптацм I дистрофнному процеа, викликаному оперативним шляхом.
8. Установити. особливост! обм!ну речовин у хворих остеохондрозом I гострим ¡нфарктом м!окарда за результатами визначення концентрац1й л1п1д!в та лт!дних фргакцШ, продукта перекисного окисления л!п1д1в (ПОЛ), кальцитоЫна, паратгормона, калыию, хондро!т1нсульфат1в, ф1бронектина, а також сумарно'1 антиоксидантно! активност| (АОА), активност! кислой та лужно'1 фосфатаз в кров!, оц1нки функционального стану пол1морфноядер-них лейкоцит!в (ПМЯЛ). Провести "га пор1вняльну характеристику.
Дисертац1я виконана у Харкюському ¡нститут! удосконалення л!кар1в зг1дно з науково-тематичним планом (нституту (номер державно! ресстрацП теми 01. 90. 001468)
Наукова новизна.
1! Вперше на приклад! вппиву МпокЫезИ та реадаптацм розглядено ¡нтеграшю м1ж тканинами серия, судин, печ!нки, м!жхребцевих диск!в, т1л хребцт та компактно! частини стегнових к!сток за результатами досл'шження компонентш вуглеводного, лт1дного, морального обм1н1в, метабол1зму органЫного матрксу.
2. Використання комплексно! оцЫки метабол|'чних реакцШ при стреа дозволяс прогнозувати структурно-метабол1чн1 пошкодження в досл1джуваних тканинах залежно в1д (ндив!руально-типолопчних особливостей оргажзму.
3
3. Вперше на модел! "експериментального остеохондрозу" доведено вплив локальних дистроф!чних зм!н у тканинах хребту на р!вень показник!в вуглеводно.го, лЫдного, генерального обмЫв I метабол1зму орган1чного матриксу сполунно'1 тканини в судинах, серц! I печ'жц).
4. Впе| ;ие з викЬристанням пом(чених сполук ,4С-пролшу, 3Н-глюкозамжу, 3Н-глюкози, 45Са, 3Н-цАМФ й 3Н-холестерину при локальны дистроф1Чн!м пронес) показано лоруи.зння ¡нтеграцП б1охш!чних процесс м1ж тканинами орган!в к!стково-суглобово1 I серцево-судинноГ систем.
5. Вперше на п1дстав! поршняння б1ох!м!чних зм^н, одержаних на моделях ппок!нез11, реадаптацИ та "експериментального остеохондрозу" запропоновано метабол1чн! критерИ переходу в!д дистроф1Чних змж у тканин! м!жхребцевих диск1в до деструк-тивних: зниження вм!сту РНК, прол1ну, оксипролЫу, тирозину, гек-соз, гексуронових кислот, загальних л!п1д1в, фосфол!п1дш I еф1р1в холестерину, переваження р(вня продукт1а ПОП над ршнем АОА.
6. Вперше обгрунтована участь загальних патогенетичних механ!зм!в у розвитку остеохондрозу I гострого ¡нфаркту мюкарда на п1дстав1 виявлення сх)дних зм!н: накопичення в кров! хворих бета-лтопроте!д!в, кальцитоншу, зниження р1вн!в паратгормону, триглщеридш й загально! активное™ лактатдег!дрогенази (ЛДГ), а в окрем1 строки .швчення захворювань - продуклв ПОЛ, АОА, хондро1т1нсульфат1в на фон1 п!двищення функц1онально! активност1 ПМЯЛ.
7. Проведено кл!нико-методичну розробку нового способу визначення фази течП 1нфаркгу м!окарда за концентрацию хондроЖнсульфат1в в кров!.
8. Проведено мет.одичну розробку способу видтення пол¡морфноядерних лейкоцит^ з кров! людини.
Отриман! нов1 дан1 дозволяють доповнити уявлення про патогенез стресогенних захворювань (на приклад! остеохондрозу I гострого !нфаркту м^карда), що сприяе. п!двищенню ефективност! д!агностики I л!кування.
Теоретична значн1сть.
У результат! експериментальних досл!джень створено концеп-ц1ю про патогенну роль р!зноман1тних стресових вплив!в у виник-ненн! I розвитку структурно-метабол'пних порушень в к!стково-суглобов1й I серцево-судинн1й системах на р!вн1 вуглеводного, л1п!дного, мщерального обмШа та метабол!зму орган!чного матриксу сполучноТ тканини.
Встановлено, що фази стресово! реакцП в досл!джуваних тканинах (за даними б!ох!м!чних досл1джень) не зб!гаються за часом.
Встановлено, що метаболЫним проявлениям ЫтеграцЦ м!ж к1стково-суглобовою та серцево-судинною системами е змжи 1снуючих в норм1 сп(вв!дношень м1ж метабол(тами та 1нш!ми показниками р!зних стор1н обм!ну речовин:
переважне використання для покриття енерговитрат в ход! реал!зацП стресово! реакцП тканинами судин й стегнових к!сток -вуглевод!в, печ!нки - л!п!д!в, а тканинами т!л хребц!в ! м!жхребцевих диск1в - позачергово углевод1в. I л!п1д1в аалежно в!д фази стресово! реакцн на фон! збереження запас!в цих речовин у тканин! м!окарда;
р(эний ступ!нь накопичення оксипрол!на у дост!джуваних тканинах при стрес1 на фон1 однаково ¡нтенсивного використання його попередника пролЫа, а, можливо, ! "конкуренцп" за цей метабол!т; '
зм1ни сп1вв1дношення кат!он!в (Са++, Си++, Ре++, Мд++) м!ж Досл1джуваними тканинами при стрес! пор!вняльно з нормою та 1нщ.
В основ! виникнення структурно-метабол!чних порушень у тканинах орган!в кожно") з досл!джуааних систем лолягае порушення ¡нтеграцн 61ох1м1чних процеЫв не т!льки в окрем!й тканин), але й м)ж тканинами р)зних систем.
Зг1дно з комплексною оценкою стану метабопЫму, в уЫх досл!джувани : тканинах у в!дпов!дь на стресовий вплив одноча-сово актив!зуються ¡деструюивж, I репаративн! процеси, сп!вв1д-ношення штенсивност! яких змЫюеться не т!льки г залежносп в'щ факторш часу реакц!Т, в1ку, але й в!д ¡ндив'|дуально-типолог1чних особливостей оргашзму. Це дозволяе прогнозувати при стрес1 структурно-метабол№н! порушення в окремих тканинах.
Показано вплив локального дистрофЫно-деструктивного процесу у хребт! на р1зн! сторони метабол!зму тканин серця, судин ! печ1нки, а також на непошкоджеж тканини к!стяка, що дозволяе теоретично обгрунтувати можлив!сть розповсюдження остеохондроза з одного в1дд!лу хребта на !нш!, можлиа1сть виникнення захворювань внутр1шн!х орган!в при остеохондроз!.
Завдяки досл!дженню м!жхребцевих диск!в при р!зних вар!-антах стресу запропоновано критерИ оц!нки переходу стану ткани-ни в!д по.чагкових дистроф(чних зм!н до деструктивних: зниження вм!сту РНК, прол!на, оксипрол!на, тирозина, гексоз, гексуронових кислот, загальних л1п!д'1в, фосфол!п!д1в 1 еф!р'|в холестерину, переваження р!внг продукт1в ПОЛ над р!внем АОА.
На п!дстав! сх!дних зм!н б!ох1м!чних параметра кров! хворих: накопичення бета-лтопротеш1в, кальцитонЫа, ф!бронектина, зниження р|'вн!в паратгормона, тригл1церид1в, загально! активност! ЛДГ, а в окрем1 строки вивчення - продуктов ПОЛ й АОА на фон шдвищенно! функц!онально! . активност! ПМЯЛ, - обгрунтоваж: ¡снування загальних б.1ох!м1чних механизма у патогенез остеохондроза хребта ! гострого ¡нфаркта мюкарда у людини.
_ П р а кт и чн а _ц| н н [сть
Схильжсть деяких тканин до переваження в них Деструк-тивних процес!в при стрес! в залежност! в1д ¡ндив!дуально-типолог!чних особливостей оргажзму треба враховувати при розробц! нових методш л!кування захворювань к!стково-сугло-бово! та серцево-судинно? систем у тварин I людини.
Розроблеш прост! I легко доступн! способи визначення фази течИ (нфаркту м!окарда 1 вид(лення пол!морфноядерних лейкоцит!в 3 КрОВ| людини.
Встановлення факту активацИ ПМЯЛ (за п1двищенними значеннями активностей лужно? фосфатази I м1елопероксидази, вм1сту синього д1формазану в НСТ-тест1, накопиченню в кл!тинах продукт!в ПОЛ I кальц!ю та Ыш.), а також сх!дних зм1н деяких б1ох1м1чних показниМв в кров! хворих на остеохондроз та гострий !нфаркт м!окарда (п!двищення р!вня бета-л(попроте1д1в, кальцито-нша, ф|бронектина, зниження р!вн1в паратгормона, тригл!церид1в та 1нш.), обгрунтовання наявност! загальних б1ох!м!чних механ1зм!в у !х патогенез! можуть стати основою для розробки заход1в проф1лактики них захворювань.
Результат» роботи впроваджен! I використовуються у курсах лекц(й Харк!вського державного унтерситету, Харк(вського !нституту удосконалення лжар!в (кафедри кл!нично! бюх^мм, орган(Ьац1Г охорони здоров'я, ортопед!! та травматолог!! N1) Хар<!вського зооветеринарного !нституту. Основа юложення дисертацН використовуються у науково-досл1дн1Й робот! Харк!вського ¡нституту ортопед!? ! травматологи 1м. проф. М.1. Си-тенка, Укратського ¡нституту експериментально! ветеринар!!.
Положения, то виносяться на захис'т.
На основ! анал1зу отриманих даних зформульован! I виносяться на захисг сл!дуюч! положения:
1. При стресових впливах (г!пок1нез!Т I реадаптаци) фази змЫ вм!сту оргзжчних та мЫеральних речовин у тканинах к!стяка; серця, судин I печ!нки не зб!гають за часом, але ж в!дновлення гомеостазу припускав 1снування системи 1нтеграи,и б!ох1м1чних процес1в м1ж тканинами.
2. СтупЬ.ь выявления метабол)чних реакц1й в тканинах оргажв к1стково-суглобово1, серцево-судинно! систем I печшки тварин при стрес! залежить в1д 1х в!ку й ¡ндив!дуаль.,о-типолог1чних характеристик, що дозволяе вид1лити групи ¡ндивщуумов "Ыдвищенного ризику", що характеризуются слабкою резистентною цих тканин до стресових вплив'/в.
3/ Локальн) дистроф1чно-деструктивн1 зм1ни у тканинах хребтового сегмента викликають порушення метабол!чних ре-акц1й в органах к1стково-суглобово? та серцево-судинноГ систем ' на р1гн! обм!ну вуглевод1в, л1п!д1в, кальц!я, орган1чного матрикса сполучноТ тканини. 1снують загальн! ланки 1нтеграц1Т метабол!чних процес1в I Гх порушення на р!вн! обм!ну гролша, гексоэам1н1в, цАМФ, глюкоз и, холестерина I кальц!я.
4. Перех1д в1д початкових дистроф!чних зм1н у тканинах .. хребтових сегмент1в до значних деструктивних характеризуется
змЫами ретти прказник1в обмЫу речовин: лад!нням р1вня РНК вкуп! з зниженням вм(сту оксипрол1ну, прол(ну, тирозину, гексоз, гексуронових кислот на фон! посиленого використання л1п1д(в як основного джерела енергИ, зниження р!вн1в фосфол!п!д1в I еф!р)в холестерину, лереваження 1нтенсивност1 накопичення продуктов ПОЛ над Ыдвищенням р1вня АОА.
5. Сх>дж зм!ни р1вн1в метабол!т!в та Ыш. покаэникт обм!ну речовин в кров1 хворих остеохондрозом \ гострим ¡нфарктом М10карда на фон! п!двищенно? функционально! активности пол;морфноядерних лейкоцитов дають шдставу вважати, що в роз-витку ,их захворювань беруть участь схож! б!олог!чж мехажзми.
8
Обсяг i структура роботи.
ДисертацШна робота викладена на 224 стор1нках машинописного тексту. Складаеться з вступу, огляду л1тератури, глави, що характеризуе постановку експерименту, матер!ал ¡ методи досл1джень, 7 глав експериментальних досл!джець, заключения, bhchobkíb по робот| i додатка (вид1леного окремим томом). Дисертат'я м1стить 265 малюнк1в i 67 таблиць. Список л1тератури включае 245 в1тчизняних i 98 закордонних джерел ЫформацИ.
Апробац1я-роботи.
Матер!али роботи викладен! ¡ обговорен! у материалах VII школи по 6ioflorlI м'яз!в (м.Харк!в, 1988), обласно! конференци по стресу i патолог!! опорно-рухшного апарату (м.Харк!в, 1989), конференцН по еколог!! лгодини (м.Нальчик, 1990), ВсеукраТнсь-кому 6i0XÍMl4H0My з"!зд1 (м.Ки!в, 1991), конференц!! по структурно-функцюнальним одиницям в органах в)сцеральних систем у норм! i патолог!! (м.Харк1в, 1991), IV науковому з'"1зд! фах!вц!в по кл1н1чн!й лабораторий д!агностиц! (м.Гродно, 1992), конференц!! по проблемах прискорбного старжня (m.Kmís, 1992), конференц1ях моло-дих вчених Х1УВ (1988-1993), конференцн по б!олог!1 опорно-pyxiBHoro апарату (м.Харк)в, '1994), заЫданж ХМНТ (m.XapKiB, 1993).
Ga темою дисертацИ опубл1ковано 23 роботи, ochobhí положения роботи захищен! 3 винаходами I 11 рац. nponi зиц1ями.
2. ХАРАКТЕРИСТИКА ЕКСПЕРИМЕНТАЛЬНОГО МАТЕР1АЛУ I МЕТОДИ ДОСЛ1ДЖЕННЯ Матер1ал"ом для вивчення були тканини т1л хребц!в, . м!жхребцевих диск1в, компактно! частини стегнових kíctok, судин (аорта), серця (стЫка л1вого шлуночка) i печшки,. а також сироватка кров! "810 безпор!дних б1лих щур1в-самц!в 3- i 129
маячного в1ку, що знаходились на стандартному pauioHi Bieapia Харктського 1нституту удосконалення лекарш. Догляд за теаринами I ix эабж здтснювали зг¡дно з правилами проведения po6¡T з використанням експериментальних тварин.
Досл!джували стан пол^морфноядерних лейкоците • -
(нейтрофил1в/ i деяк! метабол1чн1 параметры (бета-лтопротещи, фШронектин, паратгормон, кальцитон1н, кальцш, д[енов1 кон'югати, малоновий д!альдег!д, АО А, ЛДГ) у сироватц! кров! 32 хворих поперековим остеохондрозом i 45 хворих .гострим ¡нфарктом м!окарда 35-55 рок1в (на 1, 3, 5, 7, 10, 20 i 30 добу). Як стресов! фахтори у експепимент! використовували 30-добову rinoк!нез1ю, що часто зустр)чаеться в клжиц!, i фактор реадаптацП до нормальних умов на 1, 3, 5 добу. Ппок1нез|'ю зджснювали у сконструйованих нами кл!тках (Х1УВ, рац. проп. N 134В). Врахо-вувал1 1ндив!дуально-типолопчн1 особливост) орган1зму, як! визна-чались за допомогою тесту "в!дкрите поле" (Hall C.S., 1934; Кула-г)н Д. А. Федор1в В.К.,1969; Brodhurst P.L., 1975; Судаков К. В., 19Р9 та 1н).- За рух1вною активн1стю популяц!ю. тварин було розд1лено на 3 групп: I - !стоти -з високим р!внем рух!вно!
V с
активност) (рухом)), II - iстоти з пром)жною рух¡вною активЫстю (середньорухом!),. Ill - тварини з.низьким р!внем pyxiBHOi активност! (малорухом!) (Ведяев Ф.П., Чьорнобай Л.В.,1981; Кулаг!н Д.А., Болондинський В.К., 1986). Кров тварин одержували теля декап!тац!1. Внутр!шн1 органи i к!стяк досл1джували шляхом ун!версальних морфолог!чних метод!в (Г.Г. Автандтов, I.O. Казанцева, I.O. Круглова, 1976). Для контролю за станом 1мунно1 системи у кров! щур!в визначали циркулююч! !мунн! комплекси, к1льк!сть антит!лостворюючих кл1тин, б!лк1в комплемента 'i гемол|'з!н1в (Вяяова O.e., Ходжаева Ш. X., 1975; Стручков П.В. та !н., 1985).
Як модель дистрофино-деструктивного пронесу обрано вщшарування м1жхребцевого диску вщ т!ла хребця в хребцевому сегмент! у поперековШ облает! шляхом операцП, що приводить до порушення живлення травмованих дискт - диски з "дефектом" (Леонтьева Ф.С., 1985; Барер Ф.С. та !н., 1980). Операц1йна модель була реал1зована на тваринах старшо? в!ково! групи. Для демонстрацп порушення ¡нтеграцн бюх!м!чних проц,ес!в м1ж тканинами оргашв пстково-суглобово!, серцево-гсудинно! систем I печшки на р!вн1 вуглеводного, л!п!дного, м!нерального обмМв I метабол!зму орган!чного матриксу при "експериментальному остеохондроз!""використовували пом1чен! сполуки: '"'С-пролш, 3Н-глюкозамж, 3Н-цАМФ, 3Н-глюкозу, 45Са ! 3Н-холестерин. Заб1й щур1в зд!йснювали через 2,5 м!сяц!в пюля операцП'. Дослщжували структурно - метабол!чн! параметри не пльки тканин т!л хребцш м!жхребцевих диск!в з "дефектом " (П1, Д1) ! без нього (ПО, ДО), стегнових кусток, але I серця, судин, а також печ1нки, що не мають прямого в1дношення до деструктивного процесу у хребт!. У'зв'язку з участю печжки у регуляцИ практично вс1х процеЫв в орган!зм! (Т.Т.Берьозов Б.Ф.Коровк!н,1982) I з недостатн!м вйвченням в н!й обм!ну речовин при остеохондроз!, визначення метаболЫних параметр!в печ!нки було включено у перелк досл1джень.
Б1ох|'м!чн! досл1дження включали визначення у компактш'й частик! стегнових кюток, т!лах хребц!в, м!жхребцевих дисках, серц!, судинах. ! печ!нц! тварин таких компоненте ооган!чного матриксу сполучно) тканини, як оксипрол'м за ВХедетапп, лрол'т за ВосЮг, тирозин, гексозам1н, гексози I гексуронов! кислоти -методами, описаними Л.1. Слуцьким. Серед показник!в вуглеводного обм!ну визначали: сумарну активн1сть ЛДГ' спектрофотометр!чним методом, 1зоферменти ЛДГ - методом електрофорезу у пож'акриламщж'м гел! за Девю, лактат I п'руват -спектрофотометр!чними методами за РоИ1пд!^1 у наш1й
11
модиф!кацм (рац. npon. N1429,1964); гл!коген - антроновим
методом за В.П.Торбенко: Визначали показники л1п!дного обм!ну:
сумарн! лЫди - фосфорно-ван!л1новим методом зг1дно з
методичними рекомендации ХДУ (ред. Буланкжа Н.1.,1981),
фракц|йний склад - методом тонкошарово! хроматограф».
Продукти пе|.екисного окисления л!шд!в - д!енов1 кон"югати i
малоновий диальдепд (МДА) визначали спектрофотометр!чними
методами (Володимиров Ю. А., Арчаков A.I., 1972), сумарну анти-
оксидантну активн!сть - методом Г.Л. Клебанова з спшавт. (1968).
Екс'тракц1ю Л|'тд1в з тканин проводили за методом В.П. Вер-
болович та !н. (1984) у нашш модиф1кацП для к!сткових i хрящових
тканин (рац. про п. N2058, 1991). У тканинах оргажо kIctkobo-
суглобово! I серцево-судинно! систем тварин визначали показники
мЫерального обм1ну: кал1й i натр!й - методом полум"яно!
'фотометр!!: м!дь, зал1зо, хлор!ди, магн1й - стандартними наборами
"LACHEMA"; фосфор неоргашчний - за Ф|ске - Субарроу. Спос!б
визначення кальц1ю зд1йснювали комплексосометр!чним методом
Н.Л.Докторович у наш1й модиф!кацМ (рац.upon.N 1430, 1987).
Нукле1нов1 кислоти у тканинах визначали за Ogur. У сироватц1
кров! визначали: загальний б!лок б!уретовим методом,
л!попроте1ди (Колб В.Г., Камишн1ков В.С.,1976), хондро!т1н-
сульфати (Лапса Ю.Ю., Слуцький Л.1., 1975), активн!сть лужно!
фосфатази I I! к!сткового 1зоферменту, кисло! фосфатази
(стандартними наборами за методом Боданського),' активжсть
• креатЫкжази стандартними наборами "LACHEMA"; кальцигонж i
паратгормон - радМмунними наборами ф!рми "CIS" (Франц1я).
Пол1морфноядерн! лейкоцити (нейтроф)ли, ПМЯЛ) з кров! людини
видЫяли методом А.Л. Лобашевського (1983) у наш!й модиф1кац!|
(рац.проп.М 2022,1991). Кл!тинна суспенз1я використовувалася для
постановки н1тросиньотетразол1евого тесту за методом В. Н. Park
у молк Jj'Kauii В.В. Кл1мова ! Т.В.Кошовкжо! (1982). Адгезт ПМЯЛ
12
визначали за методом Л.А.Сгепаново! (1990). Активн1сть м1елоперокЫдази (МПО) у гранулоцитах визначали за методом Грехем-Кисля у викладенн1 1.Тодорова (1968). АктивМсть кисло! фосфатази у кл1тинах визначали методом азосполучення за Берстоу у модиф1кацИ Ю.Ф. Руденса,1.М.Буйк1са(1969), а активн1сть лужно! фосфатази за Кеплоу у модифкац!! Ю.Ф. Руденса з сп!вавт. (1969). Статистична обробка даних проводилась з використанням Т i Р- критерПв Стьюдента (Рокицький П.Ф., 1975; Лак1н Б.Я., 1984) за програмою, що була створена у лабораторп автоматизованих медичних систем "MEDSOFT" (м.Харк(в). Програг/а реал!зована у середовищ! Clipper 5.1 i функц!онуе на будь-який 1ВМ-сум1стн1й ПЕОМ.
3. РЕЗУЛЬТАТИ ДОСЛ1ДЖЕННЯ I ТХ ОБГОВОРЕННЯ
3.1 Метаболнн! зм!ни у тканинах юстково-суглобово! i серцево-судинно! систем при г1пок!незП I реадаптацЯ
При аналЫ отриманих даних про динам!ку компонент1в орган1чного матрйксу сполучно! тканини, вуглеводного, л!п'щного, м1нерального обм1н1в, вм!сту нукле1нових кислот у тканинах т!л хребщ'в, м1жхребцевих дискш, стегнових kIctok, серця, судин, а також печ!нки у ход1 вЩпов1дно! реакцИ на 30-добову г!покжез1ю i реа'даптац1ю до нормальних умов на 1, 3, 5 добу встановлено "фазовий" характер змж досл1джених б1ох!м!чних показник1в (концЕ?нтрацм оксипрол!ну, тирозину, гексозам1ну, гексуронових кислот, лактату, глкогену, л!шд1в I л1тдних фракцШ, ка;,ьц1ю, Mifli, зал!за та 1ншД. Встановлено також, що стутнь проявления та спрямован!сть цих зм1н у вивчених тканинах у ряд| випадмв - 36i-гаються, а у б!льшост1 випадк1в не зб1гаються-за часом (Мал.1-4). Окр1м того, характер зм!н досл1джених показникш метабол!зму у багатьох випадках не зб1гаеться в однакових тканинах експериментальних тварин р)зного в(ку.
Широкий спектр результате дано! роботи дозволяе зробити висновок про активну участь тканин к1стково-суглобово! системи, поруч з тканинами серця I печЫки, що вивчаються традиц!йно, у 1нтегральн1й в1дпов1д1 орган!зму на р1зн1 стресов! впливи. Так, при 30-добово! г1пок1нез1! у 3- мкячних щур1в найб!льш ¡стотн) в1дхилення ы!д контрольних значень вм1сту колагену (за оксипрол1ном) спостер!гались у тканинах стегнових к1сток (шдвищення на 70%), печ1нки (тдвищення на 30%) I м)жхребцевих диск!в (зниження на .35%). На 1 добу реадаптацИ зниження концентрацИ капагену у тканинах печ!нки I стегнових к1сток (у 2 рази) сполучалося э його накопиченням у тканинах м1жхребцевих диск1в I серця (до 130% I 200% в!д контрольного р!вня). На 3 добу реадаптац|{ найб!льш значними в!дхиленнями вщ контролю характеризувалися тканйни печ!нки (зниження у 5 раз!в) I стегнгвих к!сток (п!двищення на 80%). До 5 доби реадаптацП в тканинах м!жхребцевих диск!в, т!л хребц!в I серця 3-м!сячних щур!в було помЫено накопичення колагену (до 140, 180 I 260% в)д контрольного-р1вня), а у тканинах печ1нки, судин ! стегнових к1сток
- зниження його р1вня (до 20, 45 1 40% в1дпов1дно) (Мал.1). Це с
. сполучалося э в1дхиленнями в 1,5-3,5 рази в1д контрольних значень (як вб!к п)двищення, так I вб1к-зниження в залежност! в!д
тривалост!' . впливу) у тканинах орган1в к1стково-суглобовоТ, >
серцево-судинно! систем I печЫки концечтрацН неколагенових б1лк1в (за тирозином) I гл1копроте1д1в (за гексозам!ном). При
цьому, р)вн1 гексоэ I гексуронових кислот . в1др1знялись
/
найбЫьшими в1дхиленнями в)д контрольних значень у тканинах печ)нки, судин, м/х диск)в I стегЯэвих к)сток. При анал1з! результат^ можна лом1тити, що найб!льшими зм(нами у вм1ст! оксипролЫу характеризувались тканини, в яких у (нтактних тварин (взртих як контрольн!) його р!вень був нижче (у серц! 0,413+_0,01, печЫц; 0,24+_0,003, стегнових Метках 1,27+_0,06 г /100 г сухо!
.14
Осиблипосп дииамши депких показникш п тканинах юстково-суглобово! серцеио-судиппо? систем щур1а на 30 добу гшотнеап
та 1, 3, 5 добу реадалтацп. ( у % в1дносно контрольных значань, прийнятих за 100% )
7'
,Г 30 РА 1 РАЗ РА 6
Мал.1. Оксипрол1Н, шури, 3 ы1с.
Г 30 РА 1 РАЗ РА5
Мал. 2. Оксипрол1н, щури, 12 «Не
ГЗО РА 1 РАЗ .РА 5
Мвл.З. Гексозам1н, щури. 3 1ПС
ГЗО
РА 1
РАЗ
РАБ
Мал.4. Тироэ1н, щури, 12 М1С
ГЗО РА 1 РАЗ РА 5
Мал.5. Гликоген, шурн, 3 мае
ГЗО - гшокшезая 30 дндв РА - реадаптац¡я на 1,3,5 добу
ГЗО
РА 1
РАЗ
РА5
Мал.6. Загальн1 л!гпди, «ури.
3 и 1 с
тканини), жж у тканинах м/х диск1в (4,33+_0,07 г /100 г сухо? тканини) I судин (2,013+_0,04 г, /100 г сухо? тканини). Та ж законом|рн!сть спостер1галася при з1ставленн! результата зм!н концентраци гексозам1на.
Розглядаючи характер зм1н компонент оргажчного матриксу тканин, що р >сл1джуютЪс9, в орган!зм1 12-м1сячних щурт, можна пом1тити, що - вм1ст оксипролЫу в пор1внянн! з контролем змЫюеться у них у меншому ступен! (Мал. 2), жж у молодиНй (Мал. 1) в1ков(й груп!,щонайб1льш пом(тно у тканинах судин, стегнових к!сток Гм /п диск!в. Концентрац1я тирозину, у в1др1зненж в!д 3-м1сячних тварин, найбЫьш р1зким змженням пщвладна у тканинах судин, стегнових кюток I т!л хребцш (30100%). Вм1ст гексоза.м!ну характеризуемся високими значениями ампл^туди в!дхилення в!д контролю в ус!х тканинах тварин старшо? групи (60-300%), що досл1джуються, окр!м тканини печЫки, а р!вень гексуройових кислот - у вс!х тканинах (40-100%), окрЫ тканини т(л хребц)в.
При ан,ал1з! динамки компонентов орган!чного матриксу у двох вкових трупах ц!каво в)дзначити слЩуючу закономфшсть: змжи вм!сту оксипролжу в ус!х тканинах, що вивчаються, супроводжувалися . змЫами р!вня гексуронових кислот протилежного характеру ("дзеркальне в!дображення ") (Мал.7).
Якщо в 1нтегральн1м в1дбитт1 уявити, що зниження кон-центрац11 компоненте орган!чного матриксу сполучно"! тканини е проявлениям деструктивних, а п!двищення - репаративних про-цес!в (О.П.Тимошенко,1990), то можна вважати, що в уЫх вив-чених тканинах у в1дпов)дь на д!ю стресових фактор1в (на приклад1
30-добово? г1пок1нез11 I 5-денно1 реадаптаии? актив1руютьс?
» ■ „ . •
одночасно I деструктивна ,1 репаративж процеси. Сп1вв1Дношення ?х ¡нтенсивностей змжюеться ¡ндившуально у ; кожнш з
дослщжуваних тканин, залежить в1д eiKy, фази стресово! реакцм i типолог1чних характеристик icTOT.^f
• А '
/ \ / А
/
г \ /
i 7 v >
I / \ # / \
7 А /
100%Z__\JJ:J.___„ ;К°НТР0ЛЬ
N . Y • . ^J
I ч /\ / \
Ч / \ / V .
, \ . / \ / \ v / у Лг х
I .
• ■ \
I
I..........I.........I.........I.........I...........Мал. 7
О ГЗО РА1 РАЗ РА5
Динам!ка концентрацп оксипрол!ну -'— i гексуронових
кислот--------в сполучн!й тканин! на приклад! стегнових к!сток 3-
м!сячних щур!в при г!пок1нез!1 (ГЗО) та реадаптацИ (РА1, РАЗ, РА5) - "дзеркальне вщображення"
При характеристик вуглеводного I л1п!дного обм!н1в у тканинах, що досл!джуються, при ЗО-добово! г!пок!нез!1 I 5-деннш реадаптацм, за анал1зом динам!ки конценТрацИ гл!когена ! загальних л!п!д1в можна лом!тити, що в орган!зм! молодих тварин у тканинах судин I стегнових к!сток для покриття енерговитрат викорйстовуються, головним чином, вуглеводи, а в печ!нц! - лШди. Тканини т!л хребц!в I м/х диск!в використовують для цих ц1лей поперем1нно вуглеводи I л!п)ди, залежно в1д виду i строку впливу. Причому 1 доба реадаптацИ характеризуеться найб1льшими енергооитратами, оск!льки до даного строку у цих тканинах знижен1 концентрацИ гл!когену i загальних л1п!дт (Мал. 5, 6). Тканина серця на 1 добу реадаптацН, очевидно, покривае cboî енергодитрати за рахунок метаболШв, що поступили pz, не! з кров'ю, оск!льки у тканин! створюеться резерв гл!когену (220% вщ
,17
контрольного р!вня) i сумарних лт(д!в (210%). Найб1льшу "напругу", очевидно, дана тканина зазнае на 3 добу реадаптац!?, так як до цього строку в н>й I.ctotho знижуються резерви гл!когену (на 80%) i л!шд!в (на 170%) (Мал. 5,6). В-орган1зш тварин старшоГ групи найб!льшими енерговитратами, очевидно, характеризуются тканини суди ; (оск1лькй на протяз1 всього часу'досл)дження в них знижуеться концентрац!я глкогена в д1апазон! 100-20% i л!п!д!в -100-60%), а також м/х диск1в (але н1 на 1, як у 3-м'сячних щур!в, а на 5 добу реадаптац!!), так як у них вЩзначено зниження р1вн!в гл!когену (на 20-100%) I л!п1д1в (на 20-50%). Найменшими енерговитратами у 12-м!сячних luypie характеризувались тканини леч!нки (вм1ст гл!когена знижено т!льки на 1 добу реадаптацИ на 20%, а л!п!д!в - т1льки при 30-добово? г1пок!незн на 40%) i стегнових kIctok (запаси глкогена i лт!дш збер1гаються). У ' тканинах т1л хребц!в i м/х диск1в, як i у молодшт rpyni, слостер1гаеться поперем!нне використання вуглеводш i л1тд!в. У тканин! серця 12м!сячних щур1в, у в1дзнаку в!д 3-м!сячних, на 1 добу реадаптац(Г пор!вняно з г!пок1'нез|'ею спостер^гаеться зменшення тканхнних запас!в глЫогецу (на 10%) i л!п!д!в (на 60%), а на 3 ! 5 добу - lie нормал1зац!я. Сл!д пом!тити, що на 5 добу реадаптацИ концентрац!я глюкози I загальних л!п!д!в в кров! тварин обох в1кових груп, як !' р1вн! гл!когену I л!п!д1в у решт! дослщжених тканин, наближаеиься до контрольних значень.
Таким чином, у характер! використання глГкогенного i лЫдного депо тканийами орган!в к!стково-суглобово!, серцево-судинноГ систем i печ!нки при 30-добово! г!пок1нез!! та 5-добово! реадаптацИ спостер!гаеться р!зниця, що BKyni з тенденц!ею до
нормал1зац!Т показник!в на 5 добу реадаптац!! можна оцжити як
® . ,
проявления м!жтканинно! Ытеграц!! за енерговитратами, спрямовано? на збереження постшност1 внутр!шнього середовища органгму п1сля . реал!зацн реакци на стрес. Математичним
. 18
п!дтвердженням 1снування м1жтканинно! 1нтеграц11 на енергетичному р1вн1 обм1ну речовин I зм!ни II характеру при стрес1 е динамка коеф1ц1енту кореляцП м1ж змЫами р!вн)в однакових метабол1т1в у р)зних тканинах. Наприклад, кореляц(я м1ж накопиченням гл1когена у тканинах судин I м/х диск!а у 3-м1сячн'1х тварин зб1льшувалась в!д К=0,35 в контрол! до К=0,45; К=0,55; К=0,47 I К=0,92 в!дпов1дно на 30 добу г1пок1неэ!Т, 1,3,5 добу реадаптацП, а м!ж накопиченням л!п1д1в в1д К=-0,18 у контрол1 до К=-0,67; К=-0,25; К=0;62 I К=0,59. У 12-м1сячних щур1в кореляц!я м1ж накопиченням гл!когена в цих тканинах змГнювалась в1д К=0,45 в контрол! до К=0,05; К=0,79; К=0,25; К=0,65 в кожний строк досл1дження, а м!ж накопиченням л!п!д1в у тканинах судин I стегнових к1сток - в1д К=-0,15 у контрол! до К=-0,56; К=-0,68; К=-0,60; К=-0,30.
При розгляд! результат!в вивчення вуглеводного обм1ну у досл!джених тканинах молодих тварин були в!дзначен! "фази" накопичення лактата I п!двищення загально! активност! ЛДГ: в судинах на 1 добу реадаптацП (до 160% ! 165%), у стегнових к1стках - на 1 (до 180% I 300%) I 3 (до 120% I 260%) добу реадаптацП, в т1лах хребц!в - на 3 добу реадаптацП (до 150% I 140%), у серц| - на 3 (до 280% 1 110%) I 5 (до 200% ! 120%) добу реадаптацП, що супроводжувалось зниженням у них р1вня аеробних I шдвищенням - анаеробних ¡зофермент!в ЛДГ I св!дчило про активац!ю процеса анаеробного гл!кол1за в р1зн) строк! стресово"! реакцП. У 12-м1сячних щур!в "фази" накопичення лактату було знайдено на 1 добу реадаптацП не т!льки у тканин! судин (до 130%) 1 стегнових к!сток (до 180%), а на 3 добу реадаптацП не т1льки о серц1 (до 160%), але й в тканлн! м/х дискт (до 200% ! 160%),що,можливо, с ознакою порушення д|фузного живлення м/х диску з в!ком. Оск1льки накопичення лактату в почжц! з обох в1кових трупах супроводжувалось паджням його вмюту в
19
досл1дженйх тканинах I кров), а також незм)нно сполуналося з зниженням • вм1сту" Ырувата, було зроблено припущення про важливе значения печенки в у.сунени! надлишк1в лактата I п1рувата а р)зних тканин орган)зму.Р)эн1 строки -активацП в дослщжених тканинах анаеробного гл)кол1за (що висловлюеться в п)двищенн) р1вн!в активн^т! ЛДГ,' ЛДГ4 I ЛДГ5, лактату на фон! зниження концентрацИ п!рувата), що забезпечуе "т.ерм)нову" адаптац1ю тканин до стресу ¡.можливо, св!дчить про в1дсутн.^ть у них зб)гу стад)й стресовоТ реакцП.
При розГляд) динамки л)п)дних фракцж спостеркаеться зниження концентрацП фосфол(пщ1в у тканинах судин тварин двох в!кових труп (на 20-40%), що можна пов'язати з зм1ненням ста^у кл)тинних мембран або загально! к1лькост! кл!тин у тканин). В )нших дослШжених тканинах у 3-м1сячних тварин р1вень ' фосфпл)п)д)в зм)нювався у б1льшому ступен!: в стегнових к!стках на 1 добу реадаптацИ п!двищуаався на 40%, а на 5 добу знижався на 80%, в лечЫц! на 30 добу г)пок)незН зк )жався, а на 3 добу реадаптацИ .зростав на 40%. Було пом1чено. накопичення холестерину у тканинах судин молодих тварин (до 120-150% в!д . р!вня контролю) на фон) йога зниження у тканин) серця (до 6060%) у течП всього часу дослщження,. а також у тканинах стегнових к)сток (до 60%) I печ1нки (до 80%) в окрем! строки
л
досл)дження. Ужров) молодих тварин накопичення холестерину (до 170%) виявлялось на 30 день МпоЮнезП, а в пер)одреадаптацИ в!дбувалась нормал)зац1я його р!вня. 31ставляючи ц! дан) з динамкою концентрацП еф!р1в холестерину, можна пом)тити, що у тканинах печ!нки, серця, стегнових к1сток I кров) 3-м1сячних щур)в активЬуеться, а у тканинах судин гальмуеться процес етерифкацм
о
холестерина. Уч, можливо, слЩ розглядати як мехашзм захисту тка^ини вщ надмфного накопичення в н)й холестерину. В той час у тварик старшоГ групп це явище спостер^гаеться у тканинах тш
20
хребц1в \ стегнових юсток. Динамка концентраци вшьних жирних кислот у тканинах, що дослщжуються, I кров1 тварин свщчить про п1дсилене використання цих метаболМв в обм!н1 речовин, найб!льш ¡нтенсивному у молодих тварин.Узагальнення даних про зм!ст первинних I вторинних продукпв ПОЛ, АОА у тканинах, ию дослщжуються, дозволило в1дзначити моменти, коли накопичення ДК \ МДА сполучалось з зниженням АОА, що, мабуть, можна вважати ознакою ¡нтенсифкацн деструктивних процесш. Так, наприклад, у молодих' тварин у тканинах серця (у вс( строки дослщження), печшки (на 30 добу ппокЫезп) Г м/х диск!в (на 3 добу реадаптацп) накопичення ДК \ МДА до 140-200% в1д р|вня контролю сполучалось з зниженням АОА до 80-20%. У 12-м!сячних щур|в так! ж мбменти були в!дзначен1 у тканйн! судин (на 1 добу реадаптацп) I печжки (на 30 добу ппокшези I 5 добу реадаптацп). Окрш того, факт накопичення МДА (до 150-180%) I ДК (до 190230%) у тканиш м/х диск!в молодих тварин на 30 добу ппоктезп 1 1 добу реадаптацп, не днвлячись на пщвищену АОА (до 100-200%), \ збтьшення р!вня" МДА у тканин! отегнових кюток (до 130-230%) на фон') знижено! (до 50-30%) АОА, очевидно, також можна вважати ознакою посилення ¡нтенсивност1 деструктивних процес!в. У 12-м1сячних тварин аналопчна ситуац1я складастьсп у тканинах м/х дискш на 5 добу реадаптацп.
При характеристик динамки концентрацн нукле!нових кислот у тканинах молодих тварин, що дослщжуються, у вщповщь на вплив 30-добово! ппоктезп 5-денно"! реадаптацп у тканин! м/х диск1в спостер!гаеться одночасне зменшення вмюту ДНК (до 8570%) \ РНК (до 70-50%), що, очевидно, пояснюеться зниженням у тканин! вм!сту кл!тинних елемент!в. Збшьшення к!лькост!. РНК (до 120-220%) на фон! мало чим в!др!зняючогося вщ контрольного р1вня ДНК, пом!чене у тканинах т!л хребц!в на 30 добу г!пг ¡незП ! 1 добу реадаптацп, серця (до 160%) - на 1 добу, судин (до 16021
210%) ■ на 1 1 3 добу, а печ1нки (до 140-150%) - на 3 1 5 добу реадаптац1Г; можна", очевидно, пов'язати з прискореннямхпроцесу транскрипцП. ' . .
Сл)д вщзначити, що у 12-м1сячних тварин до найб1льш значних змш вм)сту нуклешових кислот можна в!днести "сплески" концентрац)? РНК, що спостер1гаються у тканинах стегнових к!сток на 30 добу г1лок1незН I 3 добу реадаптацп .(до 150% I 170%), а
також судин (печЫки на 3 добу реадаптац(Т (до 240% I 130%).
■у ■ '
Накопичення ДНК, характерне для тканин серця (до 160%) 1 печжки (до 130%) на 30 добу г!пок1нез1!, м/х дисмв (до 150%) I печжки (до 140%) на 1 день реадаптацм, не сполучене з эб1льшенням концентрац!? РНК, очевидно, пояснюеть.ся гальмуванням процесу транскрипцИ. До 5 доби реадаптац!Г загальний вм1ст нукле!нових кислот в дослщжених тканинах мае ' тендемц1ю до нормал1эац1?. Таким чином, п1двищення загально! к!лькост1 нуклё1нових кислот у тканинах оргашв юстково-суглобовоТ, серцево-судинно? систем I печ'чки при г!пок!незп I реадалтацН не зб!гаеться за часом й св1дчить про в!дсутн1сть зб1гу
у тканинах часу, включения генетичного апарату кл!тин; у молодих
в '■
. щурш це проявляеться на 1 добу реадаптацП у тканинах серця, судин I т!л хребц1в, на 3 добу - в печЫц!, а у 12-м1сячних тварин-на 1 добу реадаптацИ у тканинах печ1нки I м/х диск1в, на 3 добу реадаптацИ у тканинах стегнових к юто" I судин. А ктивац!я генетичного апарату при стрес!, за думкою Ф.З.Меерсона, (1985) спостеркаетьсяв умовах деф!циту енергП й являеться ланкою формування структурного баз!су адаптацн.
■ Оц1нюючи характер зм1н концентрацп кал1я I натрЫ у тканинах, що досл)джуються, сл!д водзначити,. що ампл!туда П В1дхилення в(д контрольн-их значень у вс! строки дослщження п'почжезП I реадаптацм значно менше (10-30%), шж при дослЩ :ент ¡нших компонент^ мжерального обмжу. Вщзначною
22
особлив!стю молодих тварин був п!двищений вм1ст кал1ю у тканин! стегнових к!сток (до 130% в!д р!вня контролю), що не супроводжувалось зниженням р!вня натр.1ю. Це можна пов'язати з 1нтенсиф1кац1ею Процес!в гл!кол!зу, або активац!ею с1нтезу б!лка (А.Лен!нджер, 1975). У 12-м1сячних тварин вм!ст цих елемент!в у тканинах, що дослЩжуються, був нижче контрольного р!вня або не в!др!знявся в!д нього.
До числа найб!льш эначних зм!н при розгляд1 особливостей мжерального обм!ну можна в1днести накопичення кальц!ю у тканинах судин- у тварин старшо! в1ково! групи (до 120-160%) I декальциф1кац!ю тканини стегнових к!сток тварин (до 60-60% в!д р1вня контролю) двох вжових труп при г!пок!незИ I р_адаптац1Т. Накопичення кальц1ю мало також м1сце у • тканин! т!л хребц!в молодих тварин, а зниження- у тканин! м/х диск1в тварин старшо! групи у вс! строки досл!дження. Тканина печ!нки характеризувалась зниженням його концентрац!! на 1 I 5 добу пюля виходу з 30-добово1 г!пок!нез!Т у двох в!кових трупах'(до 8050% в!д ршня контролю) ! "сплеском" (до 170%) на 3 день реадаптацНу молодшШ труп!. Фази зниження концентрац!! кальц1ю для тканини серця молодих тварин мали м1сце на 1 ! 3 добу реадаптацП (до 90-80%), а у 12-м1сячних - на. 30 добу ппокшезн (до 50%) 1 1 день реадаптац!! (до 60%).
• Як вщомо.у регуляц!! кальц!евого обм!ну у оргашзм! приймають участь паратгормон (ПТГ) ! кальцитонш (КТ). Ви-эначення 1х концентрац!! показало, що на 30 добу плокжез)! I 1 день реадаптац!! у кров! тварин обох в!кових груп вм1ст КТ переважае над р!внем ПТГ (у 2,5 - 6,5 разш), а на 3 ! 5 добу реадаптац!! - концентрац!я ПТГ переважае над дм!стом КТ (у 2,2 -4,2 рази). Проте характер зм!н р!вня кальц!ю у кров! I тканинах при цьому не у вс! строки досл1дження вЩповщае таким змжам концентрац!й остеотропних гормон1в, що, можливо, св!дчить про
23
порушення механизм^ кальщевого гомеостаза в умовах г!покжетичного стресу та його скасування.
При розгляд! концентрац!! мщ!, що входить до активного центру решти •антиоксидантних фермент!в, можна видтити там тканинж особливост!, як знижений р!вень у тканинах серця 1 тт хребц1в З-м^ячних иду'р1в (в дЩпазош 80-40%) протягом всього часу дослщження, а також пщвищений р!вень - у тканинах м/х дисмв, судин I печ!нки на 1 добу реадаптац!!, в тканин! стегнових к!сток ■ на 3 I 5 добу п!сля скасування ппок!нетичного режиму. У 12-м1сячних тварин найб!льш значн! змжи концентрацм мщ! були характеры для тканин серця (пщвищення у 3 рази), печшки (зниження до 40% \ 50%) на 1 I 3 добу реадаптацм,стегнових к!ст.ок (пщвищення до 160% I 200% на 30 добу г!покжезП I 5 добу реадаптацП, зниження до 80% на 3 день теля скасування ' ппокЫезП), що може бути пов'язано з змшенням ¡нтенсивност! використання м1д! у ход! реал!зац!! стресово! реакцп.
Оц!нюючи динам1ку вм!сту зал!за у моюдих тварин, можна пом1тити, що. у вЫх тканинах, що досл!джуються, ,окр!м тканини стегнових к1сток I печ1нки (в окрем! .строки досл1дження), р!вень зал!за був знижений (в д!апазон! 80-20% в!д контрольного значения). У 1_2-м1сячних тварин ..у тканин! т!л хребц1в спостер!гався п1к концентрац^ .зал!за на 30 добу г1пок!незн ! 3 день реадаптацн (до 120 I 130%). У тканин! м/х дисков п1к концентрац!! зал!за на 1 добу реадаптацп (до 140%) змжювався нормал1зац!ею !! р!вня на 3 1 5 день реадаптац!!. 3|ставлення отриманих даних з досл1дженнями М.В.51ленко (1989) дозволяс розглядати зм!нення концентрац!! зал1за як "зручний" механ!зм регуляцп сп!вв!дношення ¡нтенсивност! оксидантних ! антиоксидантних процес!в-, як в конкретжй тканин! в процеа адаптац!! до стреса, так I м!ж рЬними тканинами.
При розгляд! динамки концентрацм магн1я характерною особлив!стю тканин тварин молодшоТ групи було зниження його в м i с ту у тканин! лл хребцш (в д(апаз он! 80-40%), а також накопичення цього кат!ону у серц! (до 120%) i стегнових кютках (до 220%) на, 1 день реадаптац!!, у тканин! судин протягом всього часу досл!дження (в д!апазон! 120-140%). Тканчни, що досл!джуються,»у тварин старшоТ bíKoboí групи характеризувались зниженим bmíctom магн!ю (в Ытервал! 80-20%), за вийнятком тканин м/х диск!в i судин. На 30 день г!покжезм р!вень магнт в. м/х дисках був 150% в!д контрольного значения, а в судинах напротяз! всього часу досл!дження зм1нювався в .нтервал! 120140%. Оск1льки магн'й виконуе роль активатора лужно! фосфатази, його накопичення у к!стках можна пов'язати з зниженням активност! íf kíctkoboto ¡зоферменту, а у" серц!, зг!д'<о з дяними л!тератури (J. W. • Peterson, 1990; В.П.Кухарь та !н.,1991)-з зниженням активност! Na-K-АТФ-чзи.
Визначення концентрацИ хлорид!в показало, що у З-мюячних тварин накопичення цього ан!ону було характерно для тканин м/х дисюв (до 120 i 150%), судин (до 120 I 220%) _на 1 i 5 ahí реадаптац!? I печ!нки (до 120 I 130%) на 30 добу г1пок!нез1? I 3 день реадаптац!!, а у 12-м!сяч'них щур!в - у тканинах судин (до 120 -160%) I стегнових kíctok (до 130-200%) у окрем! строки дослщження.Як в!домо (N.Sperelakis, 1990) у м'язових, нервових та шших кл!тинах ¡они хлора розпод!ляються пасивно, оскЫьки для них немае !онного насосу, у. зв'язку з чим 1х концентрацИ зале.жить в[д провщност! мембрани.
Розглядаючи динам1ку концентрацИ фосфору у 'канинах, що досл!джуються, молодих тварин, можна пом!тити "сплески" лльки у тканинах печ^ки на 30 добу г!пок!незн I 3 день реадаптац!?, (до 130 ! 140%), стегнових kíctok на 30 добу г!покшез1? (до 115%) ! м/х flncKiB на 1 день реадаптац!? (до 115%) на фон! зниження у
■25 .
тканинах органе серцево-судинноТ системи. У старшш в1ков1й rpyni аналог!чними Мп1ками" концентрацн характеризувались тканини м/х дискш на 1 день реадаптац!! (до 150%)i судин на 1, 3, 5 дн1 реадаптаци (в fliana30Hi 120-150%). Мабуть, пояснения особливостей метабол1зму фосфору у тканинах печжки, серця,
• судин ч можна пов'язати з утворенням або роэщепленням макроерг!чних фосфатних зв'язк!в (L.Y. Opie, 1990), а у юстковш i хрящовт (В. П. Торбенко, Б.С. Касавжа, 1979 ; М. Н. Павлова та
• ¡н, 1989) - також i з утворенням кальц'|йфосфорних сполук, стабш1зуючих структуру сполучно! тканини, з активжстю к!сткового ¡зоферменту лужно! фосфатази.
Зютавляючи приведен! вище дат з результатами визначення коефщюнту кореляцн Mix накопиченням р>зних компонент!в орган1чного матриксу, вуглеводш, лт!д!в, кальц!ю, фосфору та ж. метаболтв у р!зних тканинах, обраних для дослщження, можна одержати непряме п!дтвердження припущення (Л.С.Пан!н,1983; Розанов О.Я. та in.,1985) про змжи характеру м1жтканинноТ кооперацн у ход| реалЬацн стресово! реакцИ на р|зних р!внях обм!ну речовмн. Наприклад, TicHOTa взаемозв'язку мЬк накопиченням г>рол!на у тканинах серця I "м/х диск1в в opraHi3Mi тварин CTapuio'i в!ково! групи, виражена величиною коефщ!енту кореляцм, яка зм!нювалась в1д К=0,927 у ¡нтактних щур!в до К=0,755; К= -0,645; К=0,00;К=-0,458 на 30 добу г!пок!нези ! 1,3,5 добу реадаптацп. Коеф!ц1ент кореляцн м1ж накопиченням кальщя у тканинах серця ! стегнових kIctok у орган!зм! тварин молодшо! BiKOBoi групи змжювався в!д К=-0,403 у 1нтактних щур!в до
' К=0,156; К=0,100; К=0,515;К=0,112, а м1ж накопиченням фосфора у тканинах серця ! м/х диск!в - в!д К=-0,116 у контрол1 до К=0,221; К=-0,506; К=-0,245; К=0,729, в!дпов!дно.
Анал!зуючи отриман! . дан! з урахуванням Ыдивщуально-типолопчних характеристик тварин, можна укласти, що змжи
26
61льшост1 показник1в досл1джених тканин орган1в kIctkobo-суглобово! i серцево-судинноТ систем рухомих (I), малорухомих (III) I тварин з пром1жною рухлив!стю (II) характеризуются однаковою спрямоважстю, але р!зною ампл!тудою в1дхилення в1д контрольних значень, що, очевидно, може бути пов'язако з наяз-н1стю "д1апазону стаб1л1зацН", що обумовлено. генетично (6.6. Сельков, 1978). Наприклад, при розгляд! обм!ну компоненте орган!чного матриксу сполучно! тканини вм!ст оксипрол!ну у серц! тварин I, II I III груп при flii г!пок(нез!Г I реадаптац|'Г п!двищувався _ (до 120%, 140% I 175% в!д р1вня контролю) I характеризувався ■ найб!лыиою ампл!тудою в!дхилення в1д контролю у рухомих uiypie, а вмГст гексуронових кислот знижувався (до 90%,75% i а0%) I характеризувався найб1льшим в1дхиленням в!д контролю у II rpyni. У тканин! м/х диск)в вм!ст гексозам!ну знижувався в ycix трьох трупах (до 75%,80% I 60%) I понад усе було виражено у малорухомих тварин. Проте, були виявлен! I якостн! розб!жност! у спрямованост) цих змГн. Так, наприклад, у сполучн1й тканин! серця лалорухомих тприн р1вень гл1копроте1д1в був знижений пор1вняно з контролем (до 80%), у той час як у И II трупах в1н п1двищувався (до 180% I 150%). Цей факт, у сполученн! з даними про .зниження р1вня протеогликан1в на фон! накопичення первиь.жх (до 140%) I вторинних (до 190%) лерекиЫв л!п1д1в при зниженнш до 80% АОА, дозволяс зробити припущення про тенденц1ю до переважегчя у тканин! серця малорухомих тварин деструктивних процес!в над в1дновлюючими. В той же час !нтенсиф|кац!я . деструктивних процеЫв I 1х переваження над репарс тивними при стрес! характер^ для тканини судин рухомих щур1в;
3.2. Метабол1чж особливост! тканин органш юстково-сугло-бово! 1 серцево-судинно! систем при дистрофнно-деструктивному процеЫ у хребл ("експериментальний остеоондроз")
Зпдно даним роботи встановлено, що локальний дистроф1ч-но-деструктивн'ий процес, що розвиваеться через 2,5 м1сяця шсля вщдшення м/х диску вщ т1ла хребця у одному поперековому сегмент храбту шляхом операци ("експериментальний остеохондроз") - приводить до ¡стотних змЫ метаболнних параметр^ тканин серця, судин I печЫки на р1зних етапах метабол1зму.
Так, при розгляд1 змЫ у тканинах серця, судин, печжки, тш хребц1в, м/х диск1в з "дефектом" I без нього, а також у компактжй тканиж стегнових к!сток тварин в умовах "експериментального остеохондрозу" на ршн| метабол1зму компонент оргажчного матриксу сполучноТ тканини, встановлено зниження у них вм1сту сумарного пролжа (до 50-20% в1д контрольного р1вня). У тканинах т1л хребцш з "дефектом" ! без нього, а також тканиж стегнових юсток спсстер1галось при цьому накопичення оксипролша (до 190%, 150% \ 200%), а у ¡нилх дослщжених тканинах (окр1м печЫки) - зниження йо.го вмюту (до 70-40%). Виникас припущення, що дистрофнно-деструктивний процес в одному хребцевому сегмент стимулюе у компактжй 1 губчаслй кютков!й тканинах синтез коллагену, у той час як у тканинах судин, серця 1 м/х дисюв з "дефектом" I без нього цей процес гальмуеться. (Остановлено також зниженнй (в ¡нтервал! 90-60%) р!вня неколагенових 61лк1в у вс/х тканинах, що дослщжуюгься, окр1м судин, глкопротещш (в ¡нтервал1 80-70%) у тканинах серця, стегнових юсток, т1л хребцш з "дефектом" I м/х диск1в-без нього. В той час, на фож зниженого вм1сту протеогл1кашв (в д!апазож 90-30%) у вс!х тканинах, окр1м • Т|л хребцш без "дефекту", спостер1галася змжа стввщношення бткового (визначеного за концентращею тирозина) I вуглеводного
28
(визначеного з? концентрацию гексоз) компоненте у комплексах неколлагенових б1лк1в э вуглеоодами: у тканин1 судин - у сторону переважання бокового компоненту, у тканинах печ!нки I м/х диск!в без "дефекту" - вуглеводного.
Визначення загального вм1сту одного з компонент ю м1нерального обм1ну - кальц1я . при "експериментальному остеохондроз!" показало накопичення його у тканинах т1л хребц!в ' з "дефектом" I без нього (до 120 I 140%), м/п диск1в з "дефектом" (до 140%) ! печ!нки (до 200%) на фон! зниження його концентрацЦ . (до 65% в!д р!вня контролю) у тканин! судин.
Сл!д в1дзначити, идо на р1вн! вуглеводного обмЫу, що оц!нюеться за вм1сттм компонентов ЛДГ-системи I концентрац!! глкогену, тканини т!л хребц!в I м /п диск1в з "дефектом" ! без нього, а також серия характеризувалися зкиженим вмятом лактата .(в д1апазон! 90-40% в1д р!вня контролю) на фон! накопичення п1рувата (в д!апазон1 130-300%) у вс!х тканинах, окр1м тканини м/х диск!в, як травмованих, так ! Ытактних. 3!ставлення. р!вн!в лактата I Ырувата пр^зодить до висновку, що у веж тканинах, що досл!джуються,эа вийнятком тканин м/х дискГв I стегнових к!сток, вм!ст п!рувата переважас. Ц! дан! у сукупност! з результатами визначення загально1 активност! ЛДГ I II ¡зофермеьг!в св'щчать про переважання . у тканинах т!л хребц!в, печ!нки I серця при "експериментальш'м остеохондроз!" аеробних процес1в, а у тк нин1 стегнових к!сток I м/х диск1в з "дефектом " - анаеробних. При розгляд! концентрац!? гл!когену у тканинах, що • досл!джуються можна пом!тити, що його запаси "виснажуютося" у тканинах серця, судин, стегнових к)сток (до 25-20% в!д контрольного р!вня) ! зменшуються у'тканинах м/х диск1в без "дефекту" (до 70%), у той час як його накопиченням характеризуются тканини т!л хребц!в (в д!апазон! 180-200%), м/х диск!в. з "дефектом" (до 24-0%) ! печшки (до 1000%).
Результаты визначення сумарних л!п(д!в показали, що 1х р!вень з61льшуеться (в д!апазон! 150-250%) в ус!х тканинах, що досл!джуються, за вийнятком м/х дискт з "дефектом", де цей показник знижуеться до 40% в!д р1вня контролю. Враховуючи приведен! данС можна сказати, що при "експериментальному ;остеох\эндрозГ у тканинах судин, серия, стегнових к!сток, тобто # орган!в, що не мають прямого вШношення до процесу у хребту интенсиф|куються енерговитрати за рахунок розщеплення ■ вуглеводш, у той час як запаси л!п1д1в поповнюются, а у '.канинах м/х диск'|в з "дефектом" енерговитрати здшснюються за рахунок розщеплення л1п!д1в, що дають б!льший енергетичний вих!д, жж вуглеводи. 10-кратне накопичення в цей пер!од глкогена у печ!нц!, на фон) зниженням р1зня л!л1Д1в I лактата у бшьшост! досл!джених тканин, очевидно, свщчить про ¡нтегральну роль печ!нки у переробц! лактата у ход! гл!конеогенеза при "експериментальному остеохондроз!" як своер!дному вар!ант! ендогенно'го стресу.
При розгляд! особливостей л!п1дного обм!ну при "експериментальному остеохондроз!", до найбтьш значних змш можна в!днести зниження вм1сту фосфол!п!д1в, (в д!апазон! 8575%) в ус!х досл1джених тканинах (окр!м печ!нки), що може бути пов'язано з змжами стану кл!тинних мембран, або з зниженням вм1сту кл1тинних елемент!в; накопичення холестерину у тканинах стегнових к!сток (до 120%) при зниженн! його вм!сту у тканинах серця (до 70%), можливо, за рахунок прискорено! е|ериф|'каци; зниження р!вня еф!р1в холестерину,'"у тканинах т1л хребц!в (в ¡нтервал! 90-80%) ! м /х диск!в (в !нтервал1 85- 70%), а також судин (до 80%), що, очевидно, можна пов'язати з п|дсиленим використанням жирних кислот (необхщних для этерифиацн) у синтез! тригл!церид!в. Накопичення тригл!церид!в (в ¡нтервал! 120-'220%) було пом1чено у вс1х тканинах, що досл^жуються, окр1м печжки, а також м/х дйск!в. Зниження вм!сту жирних кислот
30
встановлено у тканинах серця,т1л хребц1в з "дефектом" I без нього (вЧнтервал! 80-60%),а 1х накопичення - у тканинах судин I м/х диск1в двох тип(в (до 120 I 130%),
Сл1д в1дзначити, що при визначенн1 р!вн!в продуклв ПОЛ I АОА, у тканинах серця 1 судин, що не мають прямого в.дношення до дистроф1чно-деструктивного процесу у хребт1, виявлено накопичення ДК (в 1нтервал1 150-200%) I МДА (в ¡нтервал! 120220%) на фон! знижено! (в (нтервал! 60-50%) АОА, що поруч з даними про стан орган!чного матриксу, може бути ознакою, 1нтенсиф1кац!1 у цих тканинах деструктивних тенденц!й. Помнимо, що у тканин! т1л хребц1а зниження АОА (в дЫпазон! 60-30%) сполучасться з на'опиченням ДК, а у тканин! м/х диск!в з "дефектом", не дивлячись на п!двищений, р!вень АОА (190%), все одно в!дбуваеться накопичення первинни (до 170% ! вторинних , (до 400%) перекис!в л!п1д!в, що може св1дчити про посилення деструктивних процес1в у цих тКанинах в результат! порушення нормального функц!он!рування.
При ана.||з! результат!в визначення нукле1нових кислот у тканинах, що -досл1джуються,' при "екслериментальному остеохондрозГ у тканинах судин I стегнових к!сток було поЫчено накопичення РНК (до 180% I 510%) на фон! зниженого р!вня ДНК (до 92% I- 80%), що, очевидно, пояснюсться зб!льшенням швидкост! транскрипц!Г при зниженн) у тканин и кЬлысост) кл1т..нних елемент!в. У печ!нц! в!дзначалося паралельне зниження вмюту ДНК (до 60%) I РНК (до 55%),що може бути насл!дком зменшення к!лькост! кл!тин (Ф.З.Меерсрн, 1986). У лканинах т1л хребц!в з "дефектом" I м/х ди.ск!в спостер!галося зниження р'-1н!в РНК (до 50-40%) на фон! п(двищення концентрацП ДНК (в д1апазон! Ю8-120%), що, можливо, пов'язано з поручениям нормального функц!он!рування кл!тин в результат! травми хребтових сегмент!В.
Зютавлення результат^ досл1дження тканини м/х дискш при гтоюнезн, реадаптацЦ Г "експериментальному остеохондрозГ дозволило описати один з вар!антш переходу тканини в!д дистрофпних зм1н до деструктивних на р1вн1 метаболиних реакц1й. Критер^ми такого переходу можна вважати: р!зке падЫня ршня пролЫа 1 РНК у сполученн! з зниженням вм1сту колагену , I неколагенових бик'ш, гексоз I гексуронових кислот, використання лМдш як джерела енергП.зниження ршн1в фосфолт!дш, еф!рш • холестерину, лактату, п!рувату, ЛДГ1, переважання жте1 сивност1 накопичення первинних \ вторинних продуктш ПЬл над пщвищенням ршня.АОА.
Узагальнення даних паралельного визначення у тканинах оргажв кютково-суглобовоГ, серцево-судинноГ систем I печ1нки б1ох!мнних параметрш, що характеризують р!зн1 етапи обмжу речовин, при 30-до(зовоТ г1покжез!Т, реадаптацЦ 1 "експериментальному остеохондрозГ дозволило зробити висновки: 1) про активну участь.;;к1стково1 I хрящово? тканин у Ытегральн1й в1дпов1д! орган1зму,на стресов! впливи; 2) про вллив . локальних змш у цих тканинах на метабол1чн! реакцК у тканинах серця, судин I печЫки ;' 3) висказати припущення про 1снування ¡нтеграцн м1ж досл1дженими тканинами на всЬс розглядених етапах обмЫу речовин.
Використання 6 вид1в рад1оактиано! м!тки при реалнацП модел! "експериментальний остеохондроз" дало основу позначити конкретж ланки 1нтеграц11 б'юх1м1чних процеЫв. Так, наприклад, була встановлена шдвищена 1нтенсивнють поглинання пом1ченого прол1на (в д!апазон1 280-520%) на фон1 зниженого в них сумарного вм1сту ц!е? амЫокислоти. В сполученж з даними про накопичення оксипрол!ну в ткайинах стегнових кюток (до 200%), т!л хребцт з 'дефектом" (до 200%) I без нього (до 170%), зниження ршня оксипролжу в судинах (до 40%), серш (до 65%), м/х дисках (до
32
80%) отриман! результати св!дчили про 1нтенсиф!кац!ю використання прол!на тканинами т1л хребц!в I стегнових кюток - на посилення синтезу колагену, а у 1нших тканинах (окр1м пеу!нки) -на ¡нш1 ц!л! (можливо, енергетичн1, шляхом використання у цикл1 Кребса через утворення глутам!ново! кислоти). З^ставляюч! характер розпод!лення ЛС-прол1на м1ж тканинами, що досл(джуються, у 1нтактних тварин- з розпод!ленням його при "експериментальному остеохондроз!", можна укласти, що його метабол1зм е одною з ланок ¡нтеграцИ б1охЫ1чних процеЫв м1ж тканинами, а оск1льки тканини використовують пролш для р!зних ц1лей - не можна виключити виникнення "конкурен;них в1дносин" м!ж ними за цей ^8табол1т. I виникнення дез!нтеграц|Г на р!ен! обмЫу коллагену.
При з1ставленн1 результата розподЬпення м1чених по трШю , глюкозамШа, глюкози, цАМФ, холестерину, а також рад!оактивного кальц!ю (45Са) м!ж тканинами серця, судин, печ1нки, т1л хребц1в, м/х диск1в I стегнових к!сток !нтактних 1стот I тварин з експеримента.ьним остеохондрозом" також 'виявляються взаемообумовлен! змши обм1ну них сполук в'р!зних тканинах, як1 можна розглядати як ланки ЫтеграцН б!ох!м1чних процес!в у досл1джених тканинах.
3. 3. Особливост! метабол(зму у хворих остеохондрозом хребту I гострим 1нфарктом м!окарда
У зв'язку з тим, що одним з Лох!м1чних механ!змш ¡нтенсиф!кацИ деструктивних процеЫв у тканинах пр'" будь-якому захворюванж с активац!я перекисного окислиння лш1дш (на фон! зниження АОА), а одним з фа'кто'р!в його посилення - активац!я нейтрофЫв (пол1Морфноядерцих лейкоцитов, пмял)(ДН. Ма-янський та ¡н., 1989), що характеризуются "рест'раторним
•33
вибухом", у даЫй po6oTi провели лор!вняння функц1онального стану ПМЯЛ у хворих остеохондрозом на стад!! загострення болю i Г1М на 1, 3, 5, 7, 10, 20 i 30 день (обидва захворювання розглянут! як стресогенн!). Пор!внявши результат« проведения HCT-тесту.визначення адгез!!, активност! МПО, вмкту ДК, МДА, АОА, piBHiB' хальц1ю, загальних л!п1д1в i л!п!дних фракцШ, активност! кисло! i лужно! фосфатази встановили, що ПМЯЛ у хворих остеохондрозом i ПМ р1зних строк!в досл!дження активован!. Свщченням активацИ можуть бути виявлен! схож! змЫи ПМЯЛ: пщвищена ¡нтенсивжсть забарвлення у НСТ-тест! (до 120-220% в!д контрольного значення), пщвищений р1вень кальфю у 2-17 раз1в, активност) МПО (в д!апазон1 120-160%) i лужно! фосфатази (в д!апазож 130- 140%). До схожего можна також вщнести п|двищення у нейтроф1лах загально! активностi ЛДГ (в д1апаэош 120-150%), вм'юту сумарних л1Шд1в (в 1нтервал1 120-180%) i триглЩеридш (у 2 - 4 рази), e<pipie холестерину (в диапазон) 140160%) при остеохондроз! I у окрем! строки досл!дження при Г1М, що, очевидно, пов'язано з змженням функционального стану клкин при розвитку захворювань. Сх1дн1сть у зниженн1 АОА (до 85- 15%), концентрацК ДК (в д!апазон! 70-20%) i МДА (до 40-10%) у ПМЯЛ при даох захворюваннях, очевидно, е результатом flii ун(версального механ1зму захисту кл1тинно! мембрани в!д зруйнування власними перекисами, що вид!ляються при "респ1раторному вибусГ.
Сл1д вЩзначити, що розб!жност!, виявлен1 при з1ставленн! метаболнних характеристик нейтроф1л!в хворих (зниження до 20%, 1ндексу адгезИ 1 до 60% в!д контрольного р!вня активност) киоло! фосфатази, близьк! до контрольного значения фосфол1пщ1в i холестерина при остеохондроз'1, на в!дм1ну в1д даних ПМЯЛ при ' ПМ), очевидно, свщчать про несп(впад(ння стадШ функц!онального стану ПМЯЛ.
I 34
Аналопчний характер эм!н ом1сту метабол|т1в-маркер1в у кров1 у хворих з остеохондрозом I Г1М дав основу припустити наявн1сть эагальних патогенетичних ланок у виникненн! I роэвитку -. цих эахворювань. Так, було пом1чено накопичення атерогенних бета-л1полроте1д1а (до 130-150%), кальцитоЮна (у 2,5-;0 раз1ь), эниження р1вня сульфатованних гл1козам1ногл1кан1в хондро1т(нсульфат1в - на 3, 5 I 30 добу Г1М, а також паратгормона ' (до 80-20%), п1двищений р1вень одного э головних адгезин1в ПМЯЛ - ф1бронектина на 1, 5, 10, 20 I 30 добу (в д1апазон1 120-220%).. Отримам! реэультати св!дчать про те, що в оргашзм! хворих остеохондрозом I Г1М, можливо, мають м!сце порушення ¡нтеграцИ метабол1чних прогес!в, що . наводять до 1нтенсиф1кацН деструктивних порушень. В той же час, посилюються механ!зми,
о
що перешкоджають так!й деэ1нтеграц1ТГ наприклач, мохан1эм . захисту в!д вимивання хальц1ю з к1стковоТ тканини, або вих1д надлишку ф!бронектину у кр"в . з тканин, де в1н залучав реакц!йнозд1бн1 нойтроф1ли. Д1агностичну ц1нн1сть при виэначенн! ларактеру 1нтеграц11 обм1нних процеЫв у хворих' указаними захворюваннями мае, наприклад, визначення одного з метабол1т1в обм!ну гл1козам1ногл1кан)в в сироватц! кров1 - хондро!т1нсулйфата. При цьому, п1двищенний- його. р1вень св1дчить про розвиток у тканинах деструктивних тенденц!й, а знижений - про переваження репаративних процеЫв.
ЗАКЛЮЧЕНИЯ
ГПдводячи п1дсумки анал1зу результат1в проведених досл1джень, доц!льно роэглядати 1х у декЫькох аспектах.
Перший з них- значения отриманих даних для характеристики рол1 тканин орган1в «¡стково-суглобово? системи у. 1нтегральн1й
в!дпов!д! орган!зму на стресовий вплив на р1зних етапах метабол!зму.
Звертаючись до отриманих результате, можна побачити, що вони св!дчать на користь уявлень про активну участь тканин т!л хребц1в, стегнйвих кюток ! м/х диск1в, поруч з тканинами серия, печжк.л 1 судин, у ¡нтегральн!й в!дпов1д! орган!зму на 30-добову г!покинез!ю I реадаптац1ю тривалютю 1, 3. 5 д!б на р!вн! обм!ну компоненлв орган!чного матрйксу сполучно! тканини,
. с
вуглеводного, л!п!дного, м!нерального обмжш, нуклежовис кислот I активност! деяких фермент1в. Встановлено, що динамка вс!х компоненте, що дослщжуються, у вс!х перел1чених тканинах характ-еризуеться "фазовими змшами", . як! у ряд! випадюв зб|гають, а у б'тьшост! випадк!в- не зб!гають за часом у р1зних
V [
тканинах. Приведен! результати вкуп! з тенденц!ею до нормал1зацн бюх!м!чних показник!в до кжця перюду реадаптацн свщчать про важливу роль фактор!в регуля'ц» обмжу речовин у кожн!й тканит I необхщност! ¡снування у оргажзм! системи 1нтеграцн бюх1м1чних процес|"в м1ж тканинами для досягнення основно? цЫ адаптацн до стресу - збереження пост!йност1 внутр!шнього середовища оргаЖзму. Отриман! дан! дозволяють припустити наявн!сть таких важливих фактор!в регуляц!! обм'ту речовин, як активац!я ! гальмування процесу г1дроксил!рування пролша, використання ц!е1 амшокислоти як в синтез! колагену, так I на енергетичн! ц!л!; зм!ни спшвщношення колагена 1 протеогл!кашв у органичному матркс! сполучно! тканини, вуглеводного ! б!лкового компоненту у вуглеводно-бшкових комплексах, лактата I п!рувата, !зофермент!в ЛДГ; утил!зац!я лактата у гл!конеогенез!; етер!ф!кац!я холестерина; змши активностей ЛДГ, кисло!, а також лужно! фосфатази ! н к!сткового ¡зоферментута !н.
Другим важливим аспектом значения результатов дано! роботи с доказ впливу локальних змЫ у тканинах орган!в кютконо-
36
суглобовоТ системи (дистроф1чно - деструктивного процесу у хребцевому сегмэнтО на метабол1чн1 характеристики тканин серця, судин I печЫки як можливо! причини Ыдуцирування патологЫних зсув1в у цих органах.
Отриман! факти св!дчать про ¡стотн! енерговитрат-и 1 змь.и 61ох1м1чних показник1в у тканинах,що початково не мають прямого в!дношення до дистроф!чно-деструктивного процесу у хребт1, I ■ дають основу для пояснения випадк1в сполучення гострого 1нфаркту м1окарда I остеохондрозу з позиц1й порушення м1жтканинноТ 1нтеграц11 б1ох1м!чних процес!в.
Трет1м важливим аспектом значения результатк ц!е! роботи е визначення окремих метабол.1чних ланок ¡нтеграцИ м!ж тканинами судин, серця, печ!нки, Пл хребц1в, м1жхребцевих диск1в I стегнових к!сток. Оск1льки початковим етапом виникнення ряду захворювань, за думкою деяких дчених (П.Хочачка, Дж.Сомеро, 1988), е сама дез!нтеграц1я процес1в, що забезпечують кл1тини макроерпчними сполуками в умоаах" г!покс|Т, виникаючо? при д!Т на оргашзм ч.кстремальних фактор1в, отриман) дан1 св!дчать про кнування ланок 1нтеграц1Г та II порушення не т1льки на р!вн! обмшу вуглевод1в, л1п1д1в I цАМФ, але I на р1вн) обмшу компоненпв орган1чного матриксу сполучноТ тканини 1 кальц!я.
Четвертим важливим аспектом значения результат^ даноТ роботи е вериф(кац1я переходу початкових дистроф!чних змн у тканин! м/х диск'|в до деструктйвних на основ! анал1зу об'сктивних метабол!чних критерНв на приклад1 30-добовоГ гток)нез!Т, реадаптаци | "експериментального остеохондроза".
П'ятий аспект - встановлення вжових зм'ш стану метабол!зму у 6 видах тканин I кровI на р1вн( . вуглеводного, л1пщного, мшерального обм!н1в, орган1чного' матриксу сполучно! тканини 1 нуклеЫових кислот. Встановлено, що вков! розб1жносл за б1льш1стю показник1в, обраних для' дослщження, в умовах
37
застосування ряду стресових вплив1в згладжувалися або зникали, а у деяких випадках нав'1ть придбували жшу спрямоватсть. Це вказуе на розб1жност1 у мехажзмах адаптацм до впливш на р1зних ршнях оргажзацм системи залежно в!д т)еТ метабол1чноТ ситуацм, яка виникае у' кожний пер1од онтогенезу у окремш тканиж \ 1визначдеться Т1 функц1ональним 1 метабол'1чним станом.
Шостий аспект - виявлення взаемозв'язку отриманих зм!н з ждив!дуально-ти'полопчними особливостями нервовоТ системи ■ експериментальних тварин, що дозволяе видшити груп^ тканин "пщвищеного ризику", що характеризуются найменшою ¡'ндивЦ)у-альною резистентжстю до стресового впливу.
Сьомий аспект - на основ1 сх'|диост1 I розб1жностей у стаж метабол1'чних характеристик кровI I пол/морфноядерних лейкоцит'ш (ПМЯЛ), що являються фактором посилення процеЫв ПОЛ у хворих остеохондрозом I ПМ, пере"дбачаються загальж метабол1чн1 ланки, функц!'онування яких при обох захворюваннях сприяе, з одного боку, 1нтенсиф1кацн деструктивних процеЫв, а з друго боку -захисних механ'|зм1в, що перешкоджають зруйнуванню.
' висновки"
1. Тканини оргажв к1стково-суглобово1 системи (тт хребцт, м/х дискт, стегнових юсток), поруч з тканинами оргашв серцево-судинно! системи (судин, м1окарда) I печ!нки, приймають активну участь у ¡нтегральжй В1ДПОВщ1 оргажзму на стресов! ьлливи (30-дооову ппок1незт 1 5-денну реадаптац1ю) на р1зних р1внях обмшу речовин.
2. ДинамЫа вм1сту у тканинах т!л хребц!в, м/х дискш, стегнових к1сток, серця, судин \ печжки ¡нгрид1ент1в оргашчногс матриксу сполучно! тканини, компонента вуглеводного, л1пщного
'■мжерального обмжш, нуклежових кислот, кальц1й-регулюючи> горможв при гтокжезм 1 'реадаптацП носить "фазовий" характер
за
Фази пом1чених зм!н не эб1гають за часом в р!зних тканинах одного орган1зму I у однакових тканинах у тварин р!зного В|ку. Це св!дчить про незб1гання в них стад1й стресово! реакцП та вкуп! з. тенденц!ею до нормал!зац11 показник1в до к!нця пер1оду реадаптацП може бути розглянуто як метаб0л1чне проявления 1нтеграцП м!ж органами к!стково-суглобово5, серцевс-судинно! систем I печ1нки при стрес!.
3. П1д впливом стресу (30-добовоТ г!пок!незН I реадаптацП на 1, 3, 5 добу) тканини серця, печ1нки, т!л хребц!в, стегнових к!сток, що м!стять у !нтактних тварин найменшу к1льк!сть колагену I гл!копроте!н!в, характеризувались найб!льшими змЫами цих метабол!т!в. Тканини м/х диск!в I судин, що в!др!знялись у контрол! найб!лыиим вм!стом цих речовин, при стрес! характеризувались найменшими зм1нами. Динам1ка вм!сту колагену супрсзоджувалася зм!ною концентрацН'протеогл1кян!в прогилежного характеру у вс1х тканинах, що досл!джуються.
4. В характер! використання гл1когенного I л!п!дного депо тканинами орган)в к!стково-суглобово1, серцево-судужно! систем I печ!нки при 30-добово! г1пок!нез11 1 1-5-денна1 реадаптацП спостер1гаеться р!зниця: тканини т!л хребц!в I м/х дискш 3- ! 12-м1сячних щур!в при г(пок!нез!Т I реадаптац!Т для покриття енерговитрат- використовують поперем!нно вуглеводи ! л!п1ди; в орган!зм! молодих тварин для ц)е1 ц!л!у тканинах судин ! стегнових к!сток використовуються переважно вуглеводи, у ■'канин! печЫки -л!п!ди; тканина м!окарда характеризуемся двократним накопи-енням глкогену I л!п!д1в на 1 добу реадапхацн. У тварин старшо! в!ково? групи найб!льш1 витрати вуглевод!в ! лт!д!в були в!дзначен! у тканин! суДин, а найменш! - у тканинах печ!нки ! стегнових к!сток. Цю р!зницю в сукупност! з нормал|зац!ею р1вн1в гл!когена ! лтщ1в в тканинах до к!нця перюда реадаптацП можна розглядати як проявления м1жтканинноТ ¡нтеграц!Т на енергетичному р!вн!.
'39
5. Пщвищення вшсту нукле!нових кислот, визначене у тканинах оргажв к1стково-суглобово|, серцево-судинно! систем I печжки при ппокжезн та в р|'зж строки реадаптацп, може св!дчить про незбк у них часу "вкючення" генетичного апарату кл1тин для формування структурного базюу адаптац!!: у молодих щурш це
; проявжосться на 1 добу реадаптацп у тканинах серця, судин \ тт хребщв, на 3 добу - в печЫц! (у вигляд1 накопичення РНК), а у 12-м'1сячних тварин - на 1 добу реадаптацп в тканинах печжки I м/х -диск!в (накопичення ДНК), на 3 добу реадаптацп у тканинах стегнових к!сток I судин (накопичення РНК).
6. Змження б1льшост1 б1ох!м1чних показникш у трупах тварин з рЬною рух1вною активжстю характеризуеться однаковою спрямованн!стю, але р|зною ампл'/тудою В1дхилення В1Д контрольних значень. На основ1 оц1нки комплексу бюхМчних параметрш тканин при 30-добовоГ г1пок!нез1Г I 5-денно! реадаптацп
с
встановлено, що найбшьшою схильн!стю до деструкцГ( володють тканини судин рухомих тварин, м/х дискш середньорухомих ¡стот I тканина серця малорухомих щур!в.
7. Локальний дистрофиний процес у хребл ("експери-ментальний остеохондроз") приводить до метаболЫних зсув1в не тжьки у тканинах кютково-суглобово! системи, але 1 у тканинах серця, судин I печЫки на ршж обм!ну вуглевод1в, л!п1д1в, мжеральних компонента, орган!чного матриксу сполучно! тканини, вмюту нукле1нових кислот.
8. Виявлено наявн!сть загальнйх ланок ¡нтеграцп процес1в та !1 порушення м1ж тканинами к!стково-суглобово!, серцево-судинно! систем I печжки на р|'вн| обмжу пролЫа, гексоз-ам1н1в, цАМФ, глюкози, холестерину \ кальц1ю.
9. Метабол1чним критер1ем переходу тканини м/х дискш в1д ■ дистрофнних змЫень, до деструктивних можна вважати р|зке
падЫня р1вня прол!на I " РНК у сполученш з зниженням вмюту
40
колагена, нехолагенових б1лк!в, гексоз, гексуронових кислот на фон1 використання л1п1д1в як основного джерела енерг!!, зниження р!вн!в фосфол!п|д!в I еф!р!в холестерину, переважання ¡нтенсивност1 накопичення продуктщ ПОЛ над п!д8ищенням р!вня антиоксидантноТ активности
10. В ус!х тканинах, що досл!джуються у в1дпов1дь на стресовий вплив одночасно активукэться I деструктивн!, I репаративн! процеси, сп1вв1дношення 1нтенсивност! яких зм!нюеться 1ндив)дуально у кожно? тканини залежно в1д фактор1в часу, в!ку I типу нервовоТ системи.
11. Встановлено, що у хворих остеохондрозэм I гострим 1нфарктом м1окарда на оснрв! вйвчення функционального стачу пол!морфноядерних лейкоцит!в мають м1сце схож! змшення
г,
(п!двищен! значения активностей лужноТ фосфатази 1 МПО, вм!сту синього д1формазала у НСТ-тест!, накопичення у кл1тинах продуклв ПОЛ I кальцш), що с'вИчать про Тх активац1ю. На в!дм!ну вщ Г1М при остеохондроз! спостер1гаеться зниження ¡ндексу адгез1Г I р1вня активност! кисло! фосфатази, а також близьк1 до контрольного значения концентрацП фосфол!п!д1в I холестерину.
12. Схож) зм!ни р!вня метабол!т!в у кров! хворих остеохондрозом I Г1М (накопичення бета-/1!попроте!д!в I кальцитож'на, зниження р1вн!в паратгормона, тригл!цер|д!в I загально! активност! • ЛДГ, а у окрем! строки досл1дженнг Г1М зниження р!вн!в продукт!в ПОЛ, хондро!т!нсульфата, антиоксидантно! активност! ! накопичення ф|бронектину) у сполученн! з результатами досл!дження функц!онального стану ПМЯЛ дають основу вважати, що у патогенез! остеохондрозу ! Г1М беруть участь- загальн! патогенетичн! механ!зми, що приводить як до деструктивних так ! репаратйвних процес!в.
13. Установления м(сця ! рол! слолучно-тканинцих структур у ЫтегральноТ в!дпов!д! орган!зму на стресов! впливи, наявн!сть
41
загальних ланок ¡нтеграцП 61ох1м1чних реакцШ mix р1зними тканинами та ii порушення, визначення комплексу метабол^чних критернв стану тканин при дистроф!чно-деструктивних процесах дозволяе доповнити уявлення про патогенез стресогенних захворю&ань. (на приклад1 остеохондрозу i Г1М) i пщвищити "ефективнють д1агностики i л1кування.
СПИСОК РОБ1Т, НАДРУКОВАНИХ ЗА ТЕМОЮ ДИСЕРТАЦП 1. Состояние системы пируват-ЛДГ-лактат в тканях тел позвонков и межпозвонковых дисков крыс при гипокинезии в зависимости от возраста // Ортопедия, травматология и протез'ирование,1985.-К 7.-С.45-47/ соаат.Тимошенко О.П., Гусакова В.А.
2. Модифицирующее влияние кальция на эффективность лучевого воздействия при раке // Радиобиология, М.:Наука,1982.-
г
Т.XXII, вып.1.-С.108-109 / соавт. Френкель Л.А., Якимова Т.П.
3. Влияние нейрогенного стресса на содержание коллагеновых белков и уг^еводсодержащих компонентов тканей позвоночно.го сегмента белых крыс разного возраста // Сб.
'V,
Механизмы онтогенеза и их регуляция, Киев.: Наукова думка.-1987.- С.155-160/ соавт. Тимошенко О.1., Кладченко Л.А., Се. лина Ю.Ю.
4. О взаимосвязи остеохондроза позвоночника и острого инфаркта миокарда с позиций теории фагоцитоза // Ортопедия, травматология и протезирование, Харьков 1995, N 1. - С.38-42 / соавт. Хвисюк Н.И., Капрош H.A., Капрош Н.Ф.
5. Взаимосвязь сердечно-сосудистой системы и опорно-двигательного аппарата при стрессе // Ортопедия, травматология и протезирование, 1994,- N 4, С.102-103.
6. Электролитные сдвиги при лучевой терапии, рака молочной железы // Вопросы онкологии, М.: Мед-на, 1981. - Т. XXVII, N 1. -С.108-109. / соавт. Френкель Л.А., Якимова Т.П.
7. Демк1 показники метабол!зму в тканинах kIctkobo-суглобово!, серцево-судинно! систем та печ1чки г.ри "експериментальн1м остеохондроз!" // Укр. 61ох1м. журнал, 1995, N6. - С. 66-76 /сп!вавт. Кал1ман П.А, Тимошенко О.П.
8. Устройство для исследования активности животного. A.c.N 891067 // Бюлл.Открытия, изобретения, пром. образцы, товарн. 3H..1987.-N 45. ■
9. Устройство для проведения хронических экспериментов по иммобилизации животных // Рац. предл. УИУВ, Харьков.- 1987.-Удостовер. ,\| 1348.
10. Спос1б вид1ленння пол!морфноядерних лойкоцит!в з кров' людини. Заявка на винах1д N 94086582 в1д 8.08.1994 / сгнвавт. Капрош H.A., Шустваль Н.Ф., К1ношенко К.Ю.
11. Спос1б визначення фази теч11 1нфаркту м1окарда. Заявка на винах1д N 9"086740 в1д 23.08.1994 / cniaaBT. Капрош H.A.
12. Моделирование репаративного' остеогенеза в эксперименте // Сб. Современные проблемы остеогенеза.-Йошкар-Ола: Марийский .госуниверситет, 1982.-С.245- 247 /соавт.
.Гусакова В.А.
13. Изоферменты ЛДГ в метаболизме тканей позвоночн'-ка // Тез. V Всесоюзн. биохимич. съезда, Киев, 1986.-С.293-294/ соавт. Панченко Н.И., Ланько А.И.
14. Реадаптационные возможности соединительной ткани после гипокинезии // Сб. Экология человека. Механизмы интеграции биологических систем. Проблемы адаптации, Росгов-на-Дону, 1990.-С. 124-126.
15. Ферментативная регуляция процессов гликолиза в тканях позвоночника крыс с различной подвижностью при эмоционально-
43
болевом стрессе // Тез. Симп. Молекулярные и функциональные механизмы онтогенеза, Харьков, 1987.- С. 89-90.
16. Возрастные особенности реакции тканей позвоночника на эмоционально г болевое воздействие // Тез. докл. VII школы по биологии мышц, Харьков, 1988,- 198-199.
171 Особенности углеврдного обмена в тканях позвоночника крыс с различной двигательной активностью на фоне сердечных реакций при эмоционально-болевом стрессе // Тез. конф. Стресс и патология опорно-двигательного аппарата.- Харьков,1а89.- С. 21-22.
18. К вопросу о возникновении дистрофии в соединительной ткани "различной локализации после гипокинезии // Тез. конф. Структурно-функциональные единицы и их компоненты в органах висцеральных систем в норме и патологии,- Харьков, 1991.-С.117.
19. К вопросу о методах биохимической оценки метаболической активности нейтрофилов // Тез. там же, 1991. С. 118 / соавт. Капрош Н.А.
20. Ос.обливост! реадаплйно"1 реакцн м1жхребцевих диск1в на фот змш сполучно! тканини серця п|сля г!покшезП // Тез. VI Укр.бЮх1м. зЧзду.- 1992.- 4.2.-С. 151
21. Особенности обмена кальция у больных с острым инфарктом миокарда и при остеохондрозе // Тез.IV науч. съезда специалистов по клинической лабораторной диагностике республики Беларусь, Гродно.-1992.- С. 77-78 / соавт. Панченко Н.И., Капрош Н.А.
22. Возрастные особенности липидного обмена ' в соединительной ткани после гипокинезии // Ускоренное старение, связь с возрастной патологией. Тез. конф., Киев,- 1952. С.85 / соавт. Панченко Н.И. .
23. Об общем механизме в патогенезе остеохондроза позвоночника и острого инфаркта миокарда // Тез. конф. ХИУВ к 70-летию института, Харьков.-1993.- С.18.
Князева М.В. Метаболические аспекты изменениг в тка1 ях 0praH0J костно-суставной и сердечно-сосудистой систем при стрессе. Диссертация на соискание ученой степени доктора биологических наук по специальности 03.00.04 - биохимия, Харьковский институт усовершенствования врачей, Харьков, 1994.
Защищается 20 научных работ и 3 изобретения, которые содержат исследования, свидетельствующие об активном участии соединительной ткани различной' локализации в интегральном ответе организма на стрессовое воздействие, на разных этапах обмена веществ. Доказано существование общих метаболическиу звеньев интеграции между тканями костно-суставной и сердечнососудистой систем. Установлено также, что локальный дистрофически-дестр'уктивный процесс в позвоночнике при "экспериментальном остеохондрозе" вызывает существенные метаболические изменения в тканях сердца и'сосудов. Результаты изучения функционального состояния полиморфноядерных лейкоцитов' и некоторых метаболитов 1.рови больных остеохондрозом и инфарктом миокарда дают основание считать, что в патогенезе этих заболеваний участвуют общие механизмы, приводящие как к деструктивным, так и репаративным процессам.
Knyazeva M.V. Metabolic aspects of changes in the organs of bone-reumatic and cardiovasculaire system tissues while stress. Thesis for getting a Doctor of Biology Degree, speciality 03.00.C4 -biochemistry,Kharkov Institute of' Doctor Improvement, Kharkov 1995.
?.0 scientific works and 3 inventions which contain researches testifying connective tissue of diverse localization active participation
metabolism arej being defended.The existence of generel metabolic
bone-reumatic and cardiovasculaire system integration links between tissues'has been proved. It is also determined that local distrophic-degenerative process in a case "experimental osteochondrosis" causes considerable metabolic changes in heart and vessel tissues. The results of study of polymorphnonuclear leucocyte and some blood metabolites functional state of .osteochondrosis and myocardium infarct patients give reasons to think that eneral mechanisms leading either to destructive or to reparation processes take part in pathogenesis of these deseases.
КЛЮЧОВ1 СЛОВА
Метабол1зм, м1жхребцевий диск, т1ло хребця, стегноЕИ к1стки серцево-судинна система, ¡нтеграцЫ, стрес, дистроф1чний ороцес
in the organism's integral answer on stress on various stages of.
u

Автореферат диссертации по теме "Метаболические аспекты изменений в тканях органов костно-суставной и сердечно-сосудистой систем при стрессе."
Похожие статьи:
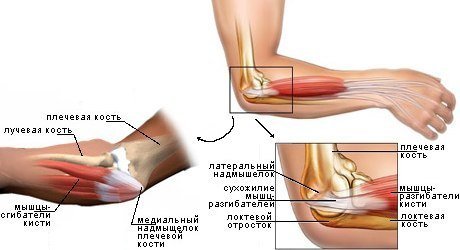 С чего начинают лечить шейный остеохондроз
Автор не ошибется, если скажет, что остеохондроз позвоночника и связанная с ним боль – одни из самых верных спутников человеческой жизни. Один человек заметил по этому поводу следующее: человечество можно разделить на три большие группы касательно их причастности к болям в спине, вызванных остеохондрозом. Первая – это те, у которых уже болело. Вторая –....
С чего начинают лечить шейный остеохондроз
Автор не ошибется, если скажет, что остеохондроз позвоночника и связанная с ним боль – одни из самых верных спутников человеческой жизни. Один человек заметил по этому поводу следующее: человечество можно разделить на три большие группы касательно их причастности к болям в спине, вызванных остеохондрозом. Первая – это те, у которых уже болело. Вторая –....
 Как лечить остеохондроз и счем
Онемение пальцев правой руки чаще всего происходит при нарушении кровоснабжения кистей и проблемах с позвоночником. Этот симптом входит также в клиническую картину многих заболеваний, которые способны приводить к очень серьезным последствиям, таким как ампутация конечности или смерть. Лечение при онемении пальцев правой руки назначается по результатам диагностики. Совокупность вызывающих гипестезию факторов можно условно разделить....
Как лечить остеохондроз и счем
Онемение пальцев правой руки чаще всего происходит при нарушении кровоснабжения кистей и проблемах с позвоночником. Этот симптом входит также в клиническую картину многих заболеваний, которые способны приводить к очень серьезным последствиям, таким как ампутация конечности или смерть. Лечение при онемении пальцев правой руки назначается по результатам диагностики. Совокупность вызывающих гипестезию факторов можно условно разделить....
 Остеохондроз цервикальная миелопатия
Грудной отдел позвоночника вместе с сочлененными ребрами выполняет функцию каркаса, защищающего жизненно важные органы от воздействия внешней среды. В силу особого строения, грудные позвонки малоподвижны, поэтому выраженная деградация межпозвоночных дисков и их разрыв наблюдается редко, а, следовательно, сильные боли при движении и нагрузке в грудном отделе спины, в отличие от шейного и поясничного отдела....
Остеохондроз цервикальная миелопатия
Грудной отдел позвоночника вместе с сочлененными ребрами выполняет функцию каркаса, защищающего жизненно важные органы от воздействия внешней среды. В силу особого строения, грудные позвонки малоподвижны, поэтому выраженная деградация межпозвоночных дисков и их разрыв наблюдается редко, а, следовательно, сильные боли при движении и нагрузке в грудном отделе спины, в отличие от шейного и поясничного отдела....
 Устранить боль остеохондроз
Занимаясь в тренажёрном зале можно не только сбросить вес или набрать необходимую мышечную массу, но и улучшить состояние мышц и связок позвоночника. Развитая мускулатура спины снимает излишнюю нагрузку с позвоночника, замедляя процесс старения этого органа, а активное кровообращение, которое возникает во время тренировки, способствует восстановлению межпозвоночных дисков. Правила проведения тренировки для оздоровления позвоночника 1....
Устранить боль остеохондроз
Занимаясь в тренажёрном зале можно не только сбросить вес или набрать необходимую мышечную массу, но и улучшить состояние мышц и связок позвоночника. Развитая мускулатура спины снимает излишнюю нагрузку с позвоночника, замедляя процесс старения этого органа, а активное кровообращение, которое возникает во время тренировки, способствует восстановлению межпозвоночных дисков. Правила проведения тренировки для оздоровления позвоночника 1....
 Остеохондроз доктор ру какие лекарства пить
Заболевания позвоночника Спондилолистез — это состояние, при котором отмечается дефект части позвонка, причиной которого является соскальзывание одного позвонка с другого. Спондилолистез нельзя путать со спондилолизисом, наиболее частой причиной ишемического спондилолистеза, при котором тело одного позвонка соскальзывает с другого. Стоит отметить, что истмической спондилолистез является наиболее частой причиной болей в спине у детей. Однако у....
Остеохондроз доктор ру какие лекарства пить
Заболевания позвоночника Спондилолистез — это состояние, при котором отмечается дефект части позвонка, причиной которого является соскальзывание одного позвонка с другого. Спондилолистез нельзя путать со спондилолизисом, наиболее частой причиной ишемического спондилолистеза, при котором тело одного позвонка соскальзывает с другого. Стоит отметить, что истмической спондилолистез является наиболее частой причиной болей в спине у детей. Однако у....
-
Комментариев еще нет. Будьте первым, кто оставит комментарий к этой статье!
Добавить комментарий